The raphe nuclei are traditionally considered to be the medial portion of the reticular formation, and
appear as a ridge of cells in the center and most medial portion of the brain stem.
In order from caudaltorostral, the raphe nuclei are known as the nucleus raphe obscurus, the nucleus raphe pallidus, the nucleus raphe magnus, the nucleus raphe pontis, the median raphe nucleus, dorsal raphe nucleus, caudal linear nucleus.[4] In the first systematic examination of the raphe nuclei, Taber et al.. (1960)[5] originally proposed the existence of two linear nuclei (nucleus linearis intermedius and nucleus linearis rostralis). This study was published before techniques enabling the visualization of serotonin or the enzymes participating in its synthesis had been developed, as first demonstrated by Dahlström and Fuxe in 1964.[6] Later, it was revealed that of these two nuclei, only the former (nucleus linearis intermedius, now known as the caudal linear nucleus), proved to contain serotonin-producing neurons,[7] though both of them contain dopaminergic neurons.[8]
In some works (e.g.[9]), researchers have grouped the nuclei lineares into one nucleus, the nucleus linearis, shrinking the number of raphe to seven, e.g.,
NeuroNames makes the following ordering:[10]
The Latin names commonly used for most of these nuclei are grammatically and orthographically incorrect. Latin grammar would require to use the genitive caseraphes ('of the seam') instead of the nominative caseraphe ('seam') in these Latin expressions. The main authority in anatomical names, Terminologia Anatomica uses for example nucleus raphes magnus[11] instead of the grammatically incorrect nucleus raphe magnus. The spelling raphe/raphes however can also be contested as numerous sources[12][13][14] indicate that raphe is an incorrect Latin rendering of the Ancient Greek word ῥαφή as the initial letter rho with rough breathing (spiritus asper) is normally rendered as rh in Latin.[12] The edition of the Nomina Anatomica that was ratified in Jena in 1935 used rhaphe instead of raphe.[15][16]
These nuclei interact with almost every pertinent portion of the brain, but only a few of them have specifically independent interaction. These select nuclei are discussed as follows.
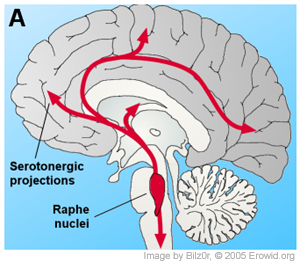
Overall, the caudal raphe nuclei, including the nucleus raphe magnus, nucleus raphe pallidus and nucleus raphe obscurus, all project towards the spinal cord and brain stem. The more-rostral nuclei, including the nucleus raphe pontis, nucleus centralis superior (also called median raphe nucleus, mRN) and nucleus raphe dorsalis (dRN) project towards the brain areas of higher function [17]
The numerous projections from the mRN and dRN to key brain structures make serotonergic system fundamental in regulating brain homeostasis. However, studies also show feedback loops from numerous areas of the brain controlling the serotonergic neurons located in the nucleus raphe dorsalis, including the orbital cortex, cingulate cortex, medial preoptic area, lateral preoptic area, and several areas of the hypothalamus. The connection between these areas, particularly between the nucleus raphe dorsalis and the orbital cortices, is thought to have influences on depression and obsessive compulsive disorder prognosis.[18]
The raphe nuclei have a vast impact upon the central nervous system.
Many of the neurons in the nuclei (but not the majority) are serotonergic; i.e., contain serotonin, a type of monoamine neurotransmitter and are modulated through fibrous pathways in the midbrain.[19]
Projections from the raphe nuclei also terminate in the dorsal horn of spinal gray matter where they regulate the release of enkephalins, which inhibit pain sensation.
The raphe nuclei provide feedback to the suprachiasmatic nuclei (SCN), thus contributing in circadian rhythms in animals. The SCN transmits to the raphe nuclei via the dorsomedial hypothalamic nucleus altering serotonin levels for sleep/wake states. The raphe nuclei will then transmit feedback to the SCN about the animal's vigilance and levels of alertness. This reciprocal feedback between the two structures provides an adaptable yet stable basis of circadian rhythms.[20]
A large increase in sympathetic nerve activity was observed when an excitatory amino acid was injected into the Raphe Pallidus , resulting in both brown adipose tissue (BAT) temperature and heart rate increasing. This suggests that activation of the raphe nucleus results in an increase in sympathetic activity to the BAT.[21]
The raphe pallidus wasn't switched off using 8-OH-DPAT, which in turn reduced body temperature due to a reduced response to cold. This suggests the importance of the raphe nucleus in responding appropriately to the cold.[22]
More recent studies of the Raphe Nuclei done with rats involve the effects of ghrelin on the dorsal raphe nucleus. When administered, larger doses of ghrelin act centrally on the raphe nucleus, hippocampus, and amygdala which causes dramatic increases in food intake, memory retention, and increases in anxiety. The effects of ghrelin are seen on the raphe nucleus as soon as an hour after injection, suggesting rapid changes in the structure of the nucleus. Changes also occur after 24 hours suggesting delayed modifications as well.[23]
^Liddell HG, Scott R (1940). A Greek-English Lexicon. Oxford: Clarendon Press. revised and augmented throughout by Sir Henry Stuart Jones with the assistance of Roderick McKenzie
^Taber E, Brodal A, Walberg F (April 1960). "The raphe nuclei of the brain stem in the cat. I. Normal topography and cytoarchitecture and general discussion". The Journal of Comparative Neurology. 114 (2): 161–87. doi:10.1002/cne.901140205. PMID13836517.
^Dahlstroem A, Fuxe K (1964). "Evidence for the Existence of Monoamine-Containing Neurons in the Central Nervous System. I. Demonstration of Monoamines in the Cell Bodies of Brain Stem Neurons". Acta Physiologica Scandinavica. Supplementum. 232 (Suppl): SUPPL 232:1–55. PMID14229500.
^Halliday GM, Törk I (April 1989). "Serotonin-like immunoreactive cells and fibres in the rat ventromedial mesencephalic tegmentum". Brain Research Bulletin. 22 (4): 725–35. doi:10.1016/0361-9230(89)90092-0. PMID2736398.
^ abHyrtl, J. (1880). Onomatologia Anatomica. Geschichte und Kritik der anatomischen Sprache der Gegenwart. Wien: Wilhelm Braumüller. K.K. Hof- und Universitätsbuchhändler.
^Foster FD (1891–1893). An illustrated medical dictionary. Being a dictionary of the technical terms used by writers on medicine and the collateral sciences, in the Latin, English, French, and German languages. New York: D. Appleton and Company.
^Triepel H (1910). Die anatomischen Namen. Ihre Ableitung und Aussprache. Mit einem Anhang: Biographische Notizen. (Dritte Auflage). Wiesbaden: Verlag J.F. Bergmann.
^Kopsch F (1941). Die Nomina anatomica des Jahres 1895 (B.N.A.) nach der Buchstabenreihe geordnet und gegenübergestellt den Nomina anatomica des Jahres 1935 (I.N.A.) (3. Auflage). Leipzig: Georg Thieme Verlag.
^Stieve H (1949). Nomina Anatomica. Zusammengestellt von der im Jahre 1923 gewählten Nomenklatur-Kommission, unter Berücksichtigung der Vorschläge der Mitglieder der Anatomischen Gesellschaft, der Anatomical Society of Great Britain and Ireland, sowie der American Association of Anatomists, überprüft und durch Beschluß der Anatomischen Gesellschaft auf der Tagung in Jena 1935 endgültig angenommen. (Vierte Auflage). Jena: Verlag Gustav Fischer.
^Peyron C, Petit JM, Rampon C, Jouvet M, Luppi PH (January 1998). "Forebrain afferents to the rat dorsal raphe nucleus demonstrated by retrograde and anterograde tracing methods". Neuroscience. 82 (2): 443–68. doi:10.1016/s0306-4522(97)00268-6. PMID9466453.
^Azmitia EC, Segal M (June 1978). "An autoradiographic analysis of the differential ascending projections of the dorsal and median raphe nuclei in the rat". The Journal of Comparative Neurology. 179 (3): 641–67. doi:10.1002/cne.901790311. PMID565370.
^Carlini VP, Varas MM, Cragnolini AB, Schiöth HB, Scimonelli TN, de Barioglio SR (January 2004). "Differential role of the hippocampus, amygdala, and dorsal raphe nucleus in regulating feeding, memory, and anxiety-like behavioral responses to ghrelin". Biochemical and Biophysical Research Communications. 313 (3): 635–41. doi:10.1016/j.bbrc.2003.11.150. PMID14697239.
Sari Y (October 2004). "Serotonin1B receptors: from protein to physiological function and behavior". Neuroscience and Biobehavioral Reviews. 28 (6): 565–82. doi:10.1016/j.neubiorev.2004.08.008. PMID15527863.




{kind=link}