
染色体説
染色体説︵せんしょくたいせつ、英: chromosome theory (of inheritance)︶、遺伝の染色体説とは、遺伝の様式を染色体の性質や挙動によって説明する学説。この学説は遺伝子が染色体上にあることを示しており、現在生物学では当然の前提とされる[1]。メンデルの法則の実証、古典遺伝学の発展、分子遺伝学の基礎形成に深く関連したことで、生物学において重要である。ただしミトコンドリアDNAなど細胞核外の遺伝因子による細胞質遺伝はこれに従わない[2]。
染色体説はバッタの染色体を用いた細胞学的観察からウォルター・サットン︵Walter Sutton︶によって1902年に提唱され、トーマス・ハント・モーガン︵Thomas Hunt Morgan︶らのショウジョウバエを用いた遺伝学的研究により、1920年代ごろ確立された。

1881年に刊行された書籍に掲載されたヒト細胞の染色体︵ヴァルター ・フレミング (Walther Flemming︶: 20世紀までに古典的な細胞学は発展を遂げていた。
染色体説提唱の背景には、全ての細胞は細胞から生じるとする細胞説と、当時再発見されたばかりのメンデルの法則がある[3]。20世紀初頭、黎明期の遺伝学と、先行して発展していた細胞学の融合から、遺伝の染色体説が誕生した[3]。
メンデルの法則は1865年に報告された[4]が、長い間歴史に埋もれ、﹁再発見﹂されたのは35年後の1900年である︵詳しくはメンデルの項目を参照︶[5][6][7]。遺伝の連続性が保証される背景には細胞説があり、古典的な細胞学は、染色・観察技術の発達とともに[8]19世紀末までには発展を遂げていた[9]。またアウグスト・ヴァイスマン︵August Weismann︶は遺伝因子は生殖細胞にあるとする生殖質説を提唱しており、細胞核に遺伝物質があることが予測されていた[10][11][12]。1842年に発見された[13]染色体に関しても、続く研究でさまざまな生物種における種類や数、細胞分裂において母細胞から二つの娘細胞へと受け継がれる様子などの知見が蓄積しつつあった[11][14]︵ヴァルター・フレミング (Walther Flemming︶の項参照︶。
このように19世紀末までには染色体説の下地ができていたが、遺伝の染色体説を主張するためには、配偶子形成における染色体の挙動を示す必要があった[15]。遺伝の一過程である受精では、卵子と精子の融合によって染色体数が倍加するため、それぞれ半数の染色体を親から受け取るはずである[15]。しかし、この過程に関する知見がまだ得られていなかった[15][11]。
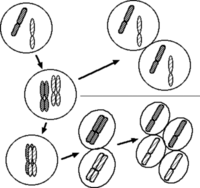
体細胞分裂と減数分裂における染色体の分配: 簡単のため一組の相同 染色体のみ示した。通常の細胞では父系と母系由来の染色体を一組ずつ持つ︵左上︶。分裂に先立って染色体の倍加が起こる︵左中︶。体細胞分裂では2倍になった染色体がそれぞれ娘細胞に受け継がれる︵右上︶。減数分裂では相同染色体が対になる︵左下︶。第1分裂で倍加したまま分配され︵下中央︶、続く第2分裂でさらに分離する︵右下︶。最終的に形成される配偶子では染色体数が半減する。
遺伝の染色体説を明確に提唱したのはウォルター・サットン︵Walter Sutton︶の1902年の論文が最初である[16]。彼はバッタの一種 Brachystola magna を用いて減数分裂の細胞学的な研究を行い[17]、配偶子形成における染色体の挙動がメンデルの法則に従うことを見いだした[16][17][18]。メンデルの法則が再発見されて間もない頃である[19][20]。
サットンはこの昆虫では染色体が大きくはっきりと観察できる利点を利用し[21][22]、配偶子形成における染色体の観察を行った。1902年の論文﹃ Brachystola magna における染色体群の形態について﹄において、配偶子形成時の細胞分裂では相同な染色体︵相同染色体︶どうしが対を作っており、これらが配偶子に一つずつ分配され、染色体数の半減、すなわち減数分裂が起こることを示した︵右図、および減数分裂の項目参照︶[17][15]。この論文の最後の段落でサットンは﹁この現象がメンデルの法則に従っており、これが遺伝の物理的基盤である可能性を示唆し、この主題について場を改めてすぐに紹介したい﹂と述べている[18][23]。そして翌年の論文﹃遺伝における染色体﹄では、この仮説をより発展させ、それぞれの染色分体がランダムに分配されることから、メンデルの法則を説明した[16]。
配偶子がもつ染色体の組み合わせは、体細胞の相同染色体対をnとしたときに2のn乗通りあり[24]、次世代における染色体の組み合わせはその2乗となる[25]。つまり4組の相同染色体をもつ場合、配偶子は42=16、次世代は162=256 通り生じる[24]。これはメンデルが交配実験で得た結果と合致する[26]︵具体例はメンデルの法則を参照︶。さらに、この論文では一つの染色体には多数の遺伝形質が存在することを予言し、またそれらは不分離だろうと述べている[注釈 1][26]。
このようにして、25歳の大学院生だったサットンによって初めて、当時知られていた細胞学の事実が、メンデルの法則という遺伝の説明に適用されたのである[20]。後に遺伝学的手法により染色体説を実証したモーガンやアルフレッド・スターティヴァント︵Alfred Sturtevant︶は﹁サットンの仮説で染色体説は既に完成していた﹂と著書や講演の中で述べている[28][16]。

ショウジョウバエの白眼変異体を用いた交配: X は交配を意味する。 黒と白のバーはいずれもX染色体を示し取り、黒のバーは赤眼の遺伝因子を、白のバーは白眼の遺伝因子をそれぞれ表す。メスはX染色体を2本、オスは1本持つ。純系赤眼のメスと白眼のオスを交配すると、次世代はオスメスともに赤眼になる。得られたメスを赤眼のオスと交配すると、次世代のメスは全て赤眼になるが、オスは半数が赤眼、半数が白眼になる。
染色体が遺伝子の担体であることを実証したのはトーマス・ハント・モーガン︵Thomas Hunt Morgan︶と彼の研究室が輩出した、いわゆるモーガン学派の研究者達である[39][40]。1904年、ウィルソンの招請によりコロンビア大学に移ったモーガン[38]は、1908年ごろからショウジョウバエを用いた遺伝学研究を始める[41]。彼らは飼育が容易で世代交代の速いショウジョウバエを用いて[42]遺伝学を発展させ、変異体の交配による遺伝学的解析と染色体の観察から、染色体説を実証していく[43]。古典遺伝学の発展と染色体説の実証は、彼らの研究成果の表裏であると言うこともできる[44]。また、染色体説を提唱したサットンを輩出した細胞学の大家ウィルソンとの交流は、モーガン学派による染色体説の発展を促進した[38]。
染色体説の背景[編集]

減数分裂における染色体の挙動と染色体説の提唱[編集]
サットンの貢献[編集]

ボヴェリの貢献[編集]
サットン以外にも染色体説を考えていた学者は少なからずいた。E. B. ウイルソン︵E. B. Wilson︶は1902年の論説﹃メンデルの遺伝の原理と生殖細胞の成熟﹄で、サットンの他にW・A・キャノンとテオドール・ボヴェリ︵Theodor Boveri︶に触れている[29][30]。コロンビア大学の学生であったキャノンは、ニューヨーク植物園の研究員時代に、サットンとは独立に、綿花を用いた妊性の実験から同様の結論に到達していた[23][31]。彼の論文は1902年に発表されている[23]。 ウィルソンが染色体説を﹁サットン-ボヴェリの染色体説﹂と呼んだことからもわかるように、ドイツの細胞学者ボヴェリも同時期に同様の構想をもっていたと言われる[19]。しかし彼はこの時期には直接的に染色体説を示唆する論文を書いていない[19]。ただしウマノカイチュウやウニを用いて研究を行っており、受精卵の染色体は卵と精子から半分ずつ由来することや、卵の細胞核を除いて受精させてもある程度まで発生が進むことを観察していた[32]。これらは染色体説を支持する観察である[33]。遺伝学による実証[編集]
モーガン学派の登場[編集]
サットンの染色体説に関する提案は、詳細なものでは最初のものであった[26]が、すぐには受け入れられなかった[11]。理由はいくつかある。まず、当時はメンデルの法則が再発見されて間もないころ[19][15]であり、遺伝子の存在に対してすら懐疑的な学者もいた[34]。染色体説を実証したモーガンも粒子遺伝をすぐには受け入れなかった[35]し、自らが証明することになる染色体説に対しても確証する結果が得られるまで長らく懐疑的だった[36]。その理由としては、サットンの仮説が実証がなされなかったことである[37]。﹁遺伝学 Genetics﹂という言葉が作られるのは1906年のことである[38]。
伴性遺伝と性染色体[編集]
モーガンは1910年に最初の突然変異体 white ︵白眼︶を発見し[45]、これを用いた交配実験の結果から遺伝子と染色体の関連性を強く示唆する結果を得た︵右図参照︶[46]。ショウジョウバエのX染色体は1908年にウィルソン研究室のネティ・スティーヴンス︵Nettie Stevens︶によって発見されており、メスはX染色体を2本、オスは1本もっていることがわかっていた[47]。純系赤眼のメスと白眼のオスを交配させると、次世代ではオスメスともに赤眼の個体が得られた。これは赤眼が白眼に対して優性なためである。ここで得られたメスと赤眼のオスを交配すると、次世代においてメスは全て赤眼だったが、オスでは赤眼と白眼が半分ずつ生まれた[注釈 2][48]。白眼の変異がX染色体上にあり、伴性遺伝するためと考えると、この現象をうまく説明できる[47][49]。この結果をより普遍化すると、染色体と遺伝との関連性、すなわち染色体説が明確に示唆されていると言える[27]。染色体遺伝学の成熟に伴う確証[編集]
1911年以降、カルビン・ブリッジス︵Calvin Bridges︶、スターティヴァント、ハーマン・マラー︵Hermann Muller︶が学生として研究室に参加[注釈 3]し、多くの突然変異体を発見しはじめる[51]。これらの変異体の交配実験の結果から、ある2つの変異体の間では、遺伝の法則の1つ﹁独立の法則﹂に従わない例が見いだされた︵[27]。既にサットンによって予測されていた[26]が、実際に得られたこの結果と考察から 連鎖︵連関ともいう‥genetic linkage︶という概念が作られた[27]。多くの変異を用いて交配実験を行った結果、それぞれの変異は4つの連鎖群に分けられたが、ショウジョウバエの染色体は4組あり、連鎖群の数と同じであった。これは遺伝子が染色体上にあることを裏付ける証拠となった[27][52]。独立の法則はそれぞれの形質が独立して遺伝するというものであり、メンデルは別々の染色体に由来する形質を観察していたため法則が成り立ったと考えられる[53]。 交配では組換え︵recombination︶が起こることも見いだされた。組換えとは同じ連鎖群に含まれる遺伝子の組み合わせが、入れ替わる現象を指す[54][27]。これに先立って、減数分裂において染色体の一部が入れ替わる交叉︵交差あるいは乗換えともいう‥crossover︶が観察されており[55]、交叉を組換えの物理的現象と考えるとうまく説明がつく[56][57]。つまり遺伝子が染色体上に線状に配置されており、組換えは交叉により染色体の一部が入れ替わったと考えられるのである[55]。また、近くにある遺伝子ほど組換えが起きにくいという推測から、スターティヴァントは線状に遺伝子の場所を特定することを発案し、遺伝子地図が作られていった[57]。組換え価の単位として用いられるcM︵センチモルガン︶は、モーガンの業績を讃えてつけられたものである[58]。 さらにブリッジスによる染色体不分離︵non-disjunction︶の発見が、染色体説を確固たるものにした[59][60]。染色体不分離とは、細胞分裂の際に染色体が均等に分かれない結果として、2個ある娘細胞のうち片方にだけ染色体が移動する現象を指す[注釈 4][61]。減数分裂においては第一段階で相同染色体が分離しない、あるいは第二段階で染色分体が分離しない現象が起こり得[注釈 5][61]、例えば性染色体では XXY やXOといった組み合わせをもった個体を生じる[62]。ブリッジスは、ショウジョウバエの交配実験において、まれな例外ではあるが、遺伝の法則に従わない形質を示す個体が得られることを発見した[63]。これらの染色体を観察すると染色体数が異常であり、染色体不分離を起こした配偶子から得られた個体であると考えられた[63][64]。これはすなわち、染色体の不分離という例外的な振る舞いが、遺伝子の例外的な遺伝と対応付けられることを示しており、染色体説の決定的な証拠となったのである[63][65]。 伴性遺伝と性染色体、連鎖群と染色体数、組換えと交叉、そして染色体不分離の実験と観察から得られた結果は、遺伝子が染色体上にあるとする考えが妥当であることを示し、染色体説を受容させるに十分であった[43]。このようにして、1920年代までには﹁遺伝子は染色体上に線状に配列している﹂ことが、事実として認められるようになった[40]。この業績により、モーガンは1933年にノーベル生理学・医学賞を受賞する[66]。 一方、モーガンがノーベル賞を受賞するよりも前、テキサス大学の T. ペインター︵Theophilus Painter︶によって多糸染色体の詳細な観察が行われた[67][68]。双翅目昆虫の幼虫にある唾液腺という組織では、細胞が細胞分裂を伴わない染色体の増幅を行うため、通常よりも太く大きい染色体を観察することができる[69]。この巨大な染色体を用いた染色体異常などの観察結果も、モーガンらの説を強く裏付けることになった[70]。染色体説以後の研究[編集]
サットンが提唱した染色体説は、モーガンの貢献により、実証された[40]。しかし一方で、遺伝子の実体は不明のままだった[40]。モーガンは、ノーベル賞受賞講演において遺伝子の物理的実体にはあまり関心が払われていないことを指摘している[67]。 遺伝子の物理的実体が明らかにされるまでには、染色体説の実証からさらに数十年の時が必要であった[71]。この間、生化学[72]や構造生物学[73]の発展に加え、細菌遺伝学[74]やファージ遺伝学[75]の発展により、染色体を構成するタンパク質と DNA のうち、DNAこそが遺伝情報の担体であることが明らかになった[76][77][78]。年表[編集]
染色体説以前[編集]
●1838-39年: シュライデン︵Matthias Jakob Schleiden︶とシュワン︵Theodor Schwann︶による細胞説の提唱[79][80]。 ●1842年: ネーゲリ︵Carl Nageli︶が染色体を発見[13]。 ●1865年: メンデル︵Gregor Mendel︶によるメンデルの法則の発表[7]。 ●1882年: フレミング (Walther Flemming︶が有糸分裂の詳細を記載[81]。 ●1890年: ヘンキング (Hermann Henking︶が カメムシの精巣細胞で奇妙な挙動示す染色体︵現在のX染色体︶を発見[82]。 ●1892年: ヴァイスマン︵August Weismann︶が生殖質説を提唱[12]。染色体説の提唱と実証[編集]
●1898年: サットン︵Walter Sutton︶が修士課程の学生としてマクラング︵Clarence McClung‥カンザス大学︶の研究室へ[83]。 ●1900年: サットンがウイルソン︵E. B. Wilson‥コロンビア大学︶の研究室へ[83]。 ●1900年: ド・フリース︵Hugo de Vries︶、チェルマク︵Erich von Tschermak︶、コレンス︵Carl Correns︶によるメンデルの法則の再発見[84]。 ●1901年: マクラングが性染色体を報告[16][82]。 ●1902年: サットンがバッタの減数分裂における染色体の挙動を報告、遺伝の染色体説を提唱[15]。 ●1903年: サットンはさらにこの説を明確に主張する論文を発表[38]。 ●1904年: モーガン︵Thomas Hunt Morgan︶がコロンビア大学へ[38]。 ●1905年: スティーヴンス︵Nettie Stevens︶が甲虫コクヌストモドキで性染色体を報告[47]。ウィルソンもハエやバッタで確認[47]。 ●1906年: ベイトソン︵William Bateson︶によって遺伝学 Genetics という言葉が作られる[38]。 ●1908年: スティーヴンズがショウジョウバエでX染色体を確認[47]。 ●1909年: ヨハンセン︵Wilhelm Johannsen︶によって遺伝子︵独: Gen, 英: Gene︶という言葉が作られる[40]。 ●1910年: モーガンの研究室でショウジョウバエの最初の突然変異体 white が発見される[85]。 ●1910-20年代: モーガンらによるショウジョウバエ遺伝学を駆使した実証[86]。 ●1914年: ボヴェリ︵Theodor Boveri︶が﹁癌の染色体説﹂[87]を提唱[32]。 ●1933年: ペインター︵Theophilus Painter︶が双翅目昆虫唾液腺の多糸染色体を詳細に解析[68]。 ●1933年: モーガンがノーベル生理学・医学賞を受賞[66]。染色体説以後[編集]
●1944年: アベリー︵Oswald Avery︶らによる肺炎双球菌の形質転換実験︵DNAが遺伝情報を担うことの実証︶[88]。 ●1952年: ハーシー︵Alfred Hershey︶らによるブレンダー実験︵ハーシーとチェイスの実験︶[89]。 ●1953年: ワトソン︵James Watson︶、クリック︵Francis Crick︶によるDNA二重らせんモデルの提唱[90]。脚注[編集]
注釈[編集]
(一)^ 実際には組換えが起こる[27][26]。
(二)^ モーガンらの実際の実験では、赤眼型のメスが2459匹、赤眼型のオスが1011匹、白眼型のオスが782匹と、想定される割合とは異なっている[48]。モーガンはこの結果を、幼虫あるいはサナギの時期の死亡率の違いによるものであるとしており、メンデルの分離の法則を実証するものであるとした[48]。
(三)^ いずれも、後に遺伝学の研究者となる[49][50]。
(四)^ 染色体説に関しては、減数分裂における不分離が取り上げられることが多いが、不分離自体は体細胞分裂でも減数分裂でも起こる現象である[61]。
(五)^ 体細胞分裂においては相同染色体が分離することはないため、染色分体の不分離のみが起こり得る[61]。
出典[編集]
(一)^ 藤川 2010, p. 44.
(二)^ 阪本, 寧男﹁3. メンデルの法則﹂﹃基礎遺伝学﹄裳華房︿21世紀への遺伝学I﹀、1995年7月20日。ISBN 4-7853-5814-9。
(三)^ ab渡辺 1998, p. 28-29.
(四)^ 渡辺 1998, p. 16.
(五)^ 渡辺 1998, p. 17.
(六)^ 渡辺 1998, p. 22.
(七)^ abデービッド・アボット 1985, p. 181-182.
(八)^ "細胞学". ブリタニカ国際大百科事典. コトバンクより2022年3月21日閲覧。
(九)^ 渡辺 1998, p. 26.
(十)^ 渡辺 1998, p. 21-22.
(11)^ abcdMartins 1999, p. 262.
(12)^ abデービッド・アボット 1985, p. 26-27.
(13)^ ab秀之, 田辺﹁染色体テリトリー: 間期核における染色体の核内配置と核高次構造に関する最近の研究﹂﹃環境変異原研究﹄第25巻第1号、2003年、13頁、doi:10.3123/jems.25.11。
(14)^ “ESP Digital Books: The Cell in Development and Inheritance, 2nd Edition” (英語). www.esp.org. p. 67. 2022年3月20日閲覧。
(15)^ abcdef渡辺 1998, p. 28.
(16)^ abcdeCrow & Crow 2002, p. 1.
(17)^ abcMartins 1999, p. 265-266.
(18)^ abSutton 1902, p. 39.
(19)^ abcdMartins 1999, p. 270.
(20)^ abMorgan 1922, p. 164.
(21)^ Crow & Crow 2002, p. 2.
(22)^ Martins 1999, p. 265.
(23)^ abcMartins 1999, p. 266.
(24)^ abSutton 1903, p. 234.
(25)^ Sutton 1903, p. 234-235.
(26)^ abcdeMartins 1999, p. 269.
(27)^ abcdef渡辺 1998, p. 43.
(28)^ Morgan 1935, p. 5.
(29)^ Wilson 1902, p. 992.
(30)^ Martins 1999, p. 267-268.
(31)^ Cannon, William Austin (1902). “A Cytological Basis for the Mendelian Laws”. Bulletin of the Torrey Botanical Club 29 (12): 661. doi:10.2307/2478693. ISSN 0040-9618.
(32)^ abデービッド・アボット 1985, p. 162-163.
(33)^ Martins 1999, p. 264.
(34)^ 渡辺 1998, p. 21-28.
(35)^ 渡辺 1998, p. 33-34.
(36)^ 渡辺 1998, p. 40-41.
(37)^ 渡辺 1998, p. 29.
(38)^ abcdef渡辺 1998, p. 30.
(39)^ 渡辺 1998, p. 44-46.
(40)^ abcde渡辺 1998, p. 50.
(41)^ 渡辺 1998, p. 34-35.
(42)^ 渡辺 1998, p. 35.
(43)^ ab渡辺 1998, p. 40-46.
(44)^ "細胞学". 日本大百科全書. コトバンクより2022年3月21日閲覧。
(45)^ 渡辺 1998, p. 37.
(46)^ 渡辺 1998, p. 39-40.
(47)^ abcde渡辺 1998, p. 40.
(48)^ abc渡辺 1998, p. 39.
(49)^ ab渡辺 1998, p. 42.
(50)^ 渡辺 1998, p. 78.
(51)^ 渡辺 1998, p. 42-43.
(52)^ Morgan 1922, p. 176-180.
(53)^ 渡辺 1998, p. 48.
(54)^ Morgan 1922, p. 185.
(55)^ abMorgan 1935, p. 7.
(56)^ Morgan 1922, p. 186.
(57)^ ab渡辺 1998, p. 43-44.
(58)^ “Morgan unit” (英語). Oxford Reference. doi:10.1093/oi/authority.20110803100209740. 2022年3月20日閲覧。
(59)^ 渡辺 1998, p. 44.
(60)^ Ganetzky, Barry; Hawley, R Scott (2016-01-01). “The Centenary of GENETICS: Bridges to the Future”. Genetics 202 (1): 15. doi:10.1534/genetics.115.180182. ISSN 1943-2631. PMC 4701082. PMID 26733664.
(61)^ abcd日本遺伝学会・遺伝学教育用語検討委員会 編﹃遺伝単改訂 遺伝学用語集対訳付き ︵﹃生物の科学遺伝﹄別冊︶﹄NTS、2021年3月15日、217-128頁。ISBN 9784860437121。
(62)^ 陽一, 三宅﹁家畜臨床分野における遺伝学、とくに染色体分析ならびにdna診断の最近の進歩﹂﹃東北家畜臨床研究会誌﹄第15巻第2号、1992年、86頁、doi:10.4190/jjvc1990.15.84。
(63)^ abcD.L.ハートル、E.W.ジョーンズ 著、布山喜章、石和貞男 訳﹃エッセンシャル 遺伝学﹄︵初版︶培風館、2005年9月8日、95-97頁。ISBN 4563077917。
(64)^ 藤川 2010, p. 29-30.
(65)^ 藤川 2010, p. 27-28.
(66)^ ab渡辺 1998, p. 46.
(67)^ ab渡辺 1998, p. 47.
(68)^ abMorgan 1935, p. 12.
(69)^ 眞路子, 茗原、益吉, 岡田﹁目で見る遺伝子発現﹂﹃化学と生物﹄第26巻第7号、1988年、437頁、doi:10.1271/kagakutoseibutsu1962.26.436。
(70)^ Morgan 1935, p. 12-14.
(71)^ 渡辺 1998, p. 34.
(72)^ 渡辺 1998, p. 70.
(73)^ 渡辺 1998, p. 90-96.
(74)^ 渡辺 1998, p. 102-103.
(75)^ 渡辺 1998, p. 120-122.
(76)^ 渡辺 1998, p. 102.
(77)^ 渡辺 1998, p. 107.
(78)^ 渡辺 1998, p. 138.
(79)^ デービッド・アボット 1985, p. 73.
(80)^ デービッド・アボット 1985, p. 74-75.
(81)^ Flemming, W (1882). Zellsubstantz, Kern und Zelltheilung. F.C.W. Vogel, Leipzig, Germany.
(82)^ ab竹中要﹁性染色体の発見史﹂︵pdf︶﹃生物の科学 遺伝﹄第11巻第4号、1957年、13-14頁。
(83)^ abCrow & Crow 2002, p. 1-2.
(84)^ デービッド・アボット 1985, p. 103-104.
(85)^ 渡辺 1998, p. 37-39.
(86)^ 渡辺 1998, p. 54.
(87)^ Victor A McKusick (1985). “Marcella O'Grady Boveri (1865-1950) and the chromosome theory of cancer” (pdf). Journal of Medical Genetics 22: 431-440.
(88)^ デービッド・アボット 1985, p. 19-20.
(89)^ 渡辺 1998, p. 122.
(90)^ デービッド・アボット 1985, p. 54-56.
参考文献[編集]
●Crow, Ernest W; Crow, James F (2002-01-01). “100 Years Ago: Walter Sutton and the Chromosome Theory of Heredity” (英語). Genetics 160 (1): 1–4. doi:10.1093/genetics/160.1.1. ISSN 1943-2631. ●Martins, L.A.-C.P. (1999-06). “Did Sutton and Boveri propose the so-called Sutton-Boveri chromosome hypothesis?”. Genetics and Molecular Biology 22 (2): 261–272. doi:10.1590/S1415-47571999000200022. ISSN 1415-4757. ●Morgan, Thomas Hunt (1922-12-01). “Croonian Lecture:—On the mechanism of heredity”. Proceedings of the Royal Society of London. Series B, Containing Papers of a Biological Character 94 (659): 162–197. doi:10.1098/rspb.1922.0053. ●Morgan, Thomas Hunt (1935). “The Relation of Genetics to Physiology and Medicine”. The Scientific Monthly 41 (1): 5–18. ISSN 0096-3771. ●Sutton, Walter S. (1903). “The Chromosomes in Heredity”. Biological Bulletin 4 (5): 231–251. doi:10.2307/1535741. ISSN 0006-3185. ●Sutton, Walter S. (1902-12). “ON THE MORPHOLOGY OF THE CHROMOSO GROUP IN BRACHYSTOLA MAGNA” (英語). The Biological Bulletin 4 (1): 24–39. doi:10.2307/1535510. ISSN 0006-3185. ●Wilson, Edmund B. (1902). “Mendel's Principles of Heredity and the Maturation of the Germ-Cells”. Science 16 (416): 991–993. ISSN 0036-8075. ●藤川, 和男﹃ショウジョウバエの再発見-基礎遺伝学への誘い-﹄︵初版︶サイエンス社︿新・生命科学ライブラリ-生物再発見 8-﹀、2010年8月10日。ISBN 9784781912585。 ●渡辺, 政隆﹃DNAの謎に挑む﹄朝日新聞社、1998年8月25日。 ●デービッド・アボット﹃世界科学者事典﹄ 1生物学者、原書房、1985年。ISBN 4-562-01609-4。関連項目[編集]
外部リンク[編集]