
星口動物
| 星口動物 | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 スジホシムシ属の一種Sipunculus robustus | |||||||||||||||||||||
| 分類 | |||||||||||||||||||||
| |||||||||||||||||||||
| 学名 | |||||||||||||||||||||
| Sipuncula Rafinesque, 1814 | |||||||||||||||||||||
| 綱 | |||||||||||||||||||||
星口動物︵ほしくちどうぶつ、ほしぐち-、Sipuncula︶は、一般にホシムシと呼ばれる蠕虫状の海産無脊椎動物。体は左右対称で、節︵体節︶に分かれていない[1]。約250種が含まれる[1]。かつては独立した門に分類されていたが、2018年現在は環形動物門の一部とされている︵詳細は環形動物を参照︶。
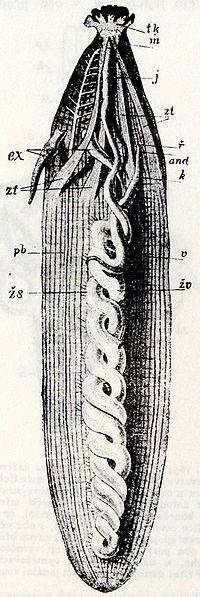
スジホシムシ属の体腔内の図
ソーセージのような体形の蠕虫。体長は3cmから10 cm程度のものが多いが、1 cmに満たない小さな種や、約50 cmに達する大型のものもいる[1]。体色はさまざまだが、くすんだ色が多い[2]。
体は体幹部と陥入吻からなる。陥入吻の先端に口があり、その周囲または背側に触手が多数ある。和名の星口とは、この触手が放射状に広がる様子に由来する[3]。体腔内に1対か2対の牽引筋があり、これによって陥入吻を体幹部に引き込むことができる[3]。種によっては吻を引き込み、体を膨らませた姿がピーナッツに似ることから、英語ではpeanut wormと呼ばれる[1]。陥入吻にはしばしば棘や鉤が並ぶ[2]。
体表はクチクラに覆われ、その表面には突起やとげがあることも多い。タテホシムシ科のものは体表の一部にクチクラが厚くなるか、石灰質が沈着してできた盾状部を持つ[1]。
クチクラの下には表皮、次いで真皮がある[1]。その内側には筋肉があり、外側に環筋層、内側に縦筋層があるほか、この2層の間に薄い斜筋層が見られることもある[1][2]。縦筋層の下には体腔膜があり、その内側が体腔である。
体腔は裂体腔性の真体腔。腹膜により仕切られているが完全ではなく、1つに繋がっている[1]。はっきりとした血管系は見られず、体の運動によって体腔液が攪拌されることが循環系の機能を果たしている[3][4]。体腔液中にはヘムエリスリンを含む赤血球があり、体表を通じて得られた酸素を輸送する[1]。ガス交換の場は種によって異なり、触手と吻の表面をともに使っているものもいるが、岩に穿孔する種や、酸素の乏しい泥などに潜る種は、触手表面だけで呼吸を行う。大型の種は体表をすべて使っている可能性がある[1]。
体腔液内には、赤血球以外にも数種類の細胞が含まれる[1]。体腔液は水力学的骨格として、圧力により体を支持したり、吻を伸ばしたりする役割も果たす[1]。
体腔の大部分は消化管が占める[3]。消化管は先端の口から後部へ伸び、後端で折り返して、体幹前端の背面にある肛門に続く。したがって消化管はU字型になり、後ろへ伸びる腸管と、後端から戻ってくる腸管は、互いに螺旋に巻きついている[3][5]。このような屈曲した消化管は、星口動物以外には、固着性の動物にしばしば見られる[2]。
腎管が1対または1本、体幹前部の腹面側にある。腎管は外腎口で体表に、内腎口で体腔内に繋がる[1][3]。窒素老廃物は腎管を通じて、アンモニアとして排泄される[1]。
小さな脳︵脳神経節︶と1本の腹神経索があり、この2つを繋ぐ食道神経環が体壁に沿って延びている[3][1]。この構造は環形動物に似ているが、ホシムシでは腹神経索が分節しないという違いがある[2]。触覚受容器は体表全体にあるが、とくに触手に多い。化学受容器は吻の背側に見られる[1]。脳神経節の背側に一対の色素杯単眼を持つ種も多い[1][2]。

フクロホシムシ属の一種。吻を伸ばし、触手を出している。

深海から採集されたサメハダホシムシ属の一種Phascolosom a turnerae
星口動物はすべて海産の底生生物である。温暖な海域に分布する種が多いが、北極海や南極海を含む世界中の海に見られ[2]、潮間帯から水深5000メートルを超える深海まで幅広い水深に生息する[1]。多くは砂のなかや石の下、海藻の根元などに棲む。サンゴ礁や沿岸域には、硬い石灰岩に穿孔するものが多い。巻貝の殻や多毛類などが作った棲管を住処とするものもいる。
柔らかい基質に巣穴を掘るときや、海藻の根元や砂利の中を動き回るときには、縦筋と環筋、吻を動かすことで蠕動する。硬い基質に穿孔するものは、体表の棘や盾状部をやすりのように使うだけでなく、分泌物によって化学的に溶かすこともしていると考えられる。タテホシムシ類は、盾状部を蓋にして巣穴の入り口を塞ぐ[1]。
タテホシムシ属のある種はスツボサンゴ属やムシノスチョウジガイ属の単体性サンゴと共生する[6]。サンゴの骨格に細い穴が開いていて、そのなかにホシムシが生活している。サンゴの骨格のなかには貝殻が見つかるので、ホシムシははじめ貝殻に棲み、その貝殻にサンゴが付着して、貝殻を覆い隠すほどに成長することで共生関係が成立すると考えられている[7][8][6]。これらの種は砂地に生息するが、サンゴは自力ではほとんど動けないので、ひっくり返ったり、砂に埋もれたりしてしまう危険がある。ホシムシがサンゴを引きずって動かすことで、サンゴは砂地を安全に利用できるようになる[6]。ホシムシの側は、サンゴが貝殻よりも大きく成長することで、自分が大きくなっても新しい住処を探す必要がなくなる[6][7]。そのほかに、サンゴは広範囲に餌を探すことができ、ホシムシはサンゴの刺胞によって外敵から身を守ることができるという利益もあると考えられる[7]。これらのことから、ホシムシとサンゴの共生は両者に利益のある相利共生と考えられる[6]。
個体によっては体表面後部に突起が確認できるが、これはこの生物に寄生する二枚貝の一種でsisoyと呼ばれ、Erycinacea科に属すると推定される[9]。

ホシムシに寄生する二枚貝︵フィリピン・マクタン島︶
採食行動には不明な点が多いが、大半の種は触手の繊毛と粘液を使って、海底表面のデトリタスを食べていると考えられる。ほかに堆積物を摂食する種や、水中の有機物を濾しとって食べる懸濁物食者もいる。石灰岩にすむ種は、吻にある鉤や棘でデトリタスをかき集めて食べる[1]。

Lecthaylusは星口動物の化石とされるが、疑問視する意見も ある。
化石記録は乏しいが、中国南西部の帽天山にあるカンブリア紀の地層から発見されたArchaeogolfingiaとCambrosipunculusの化石は星口動物のものとされている[19]。この2属は現生のフクロホシムシ科によく似ており、このグループの形態が長期間にわたって大きな変化を遂げてこなかったことを示唆している[19][20][21]。
星口動物であるとされた化石はほかにもあるが、その一部は他の分類群であることが明らかになり、残りのものも疑問がある[19]。バージェス頁岩から見つかったオットイアははじめホシムシとされたが、後にエラヒキムシであることが判明した[19][20]。シルル紀と石炭紀のLecthaylus、ジュラ紀のEpitrachysなどを星口動物とみなす意見もある[22]が、疑う意見もある[19][20]。消化管の構造が似ていることから、円錐形の殻を持つヒオリテスが星口動物に近いとする研究者もいるが、これも疑問視されている[19][20]。
星口動物自体の化石ではないが、フランスの白亜紀初期の地層から発見されたムシノスチョウジガイ類のサンゴの化石は、ホシムシと共生した痕跡を残している[8]。ほかに星口動物の生痕化石と思われる巣穴の痕跡[10]やウミユリの瘤も見つかっているが、それらが本当にホシムシの生痕かどうかは不明である[22]。

中国で食用に売られているホシムシ
食用や釣り餌用に利用されることがある。
中国福建省の料理である土筍凍は、土筍と呼ばれるホシムシの一種を煮こごりにしてタレをかけたものである[25]。山東省煙台市では、スジホシムシとユムシ類を区別せずに海腸子と呼び、ニラと一緒に炒めたものを韮菜炒海腸と称して食べる[25]。スジホシムシはベトナムでは地虫と呼ばれ、野菜と一緒に炒めて食されるほか[26]、ミクロネシアでも食用にされ[5]、日本の瀬戸内海や九州では釣り餌用に利用されている[2]。
フィリピンのマクタン島など一部地域では、スジホシムシ属のSipunculus robustusをサルポと呼び、酢と薬味を和えて生食するほか、天日干しにしたものを炙って食べることもある。フエフキダイ科の魚を釣るための餌に用いることもある[27]。砂地にあるサルポの巣穴を発見し、採集するには熟練した技術が必要で、専門的な採捕者がいる。マクタン島ではスジホシムシも採捕されるが、苦いので食用には好まれず、釣り餌や、滋養強壮のための民間薬として利用される[27]。カオハガン島ではサルポのほかに、より小さなホシムシ類がバトナン、さらに小さいものがバトレと呼ばれ、野菜と酢で和えて食べる[28]。ただし、バトナン︵Holothuria inhabilis︶とバトレ︵Holothuria arenicola︶はナマコの一種である[29]。

ホシムシ採捕の様子︵フィリピン・マクタン島︶
日本では、1920年代から1930年代ごろには広島県沿岸で採取されたスジホシムシやスジホシムシモドキが釣り餌として利用されていた記録があるが、2010年前後の西日本での調査によれば、流通しているのは日本国外から輸入されたスジホシムシのみである[30]。
特徴[編集]

生態[編集]



繁殖と発生[編集]
有性生殖[編集]
温帯では夏から初冬に繁殖する種がもっとも多いが、例外もある。繁殖行動は夜間に行われることが多い。配偶子は腎管を通じて体外に放出され、海水中で受精︵体外受精︶する。雄が先に精子を放出すると、雌が水中の精子に反応して卵を放出するという観察があるが、種によっては逆に卵が先に放出されることもある[1][10]。 ほぼすべての種が雌雄異体だが、例外的にフクロホシムシ科のNephasoma minutumは雌雄同体である。ただし自家受精はしない。この種では、受精卵は母親の巣穴のなかで保護される[10]。無性生殖[編集]
有性生殖に加えて、高い再生能力による無性生殖も行うことが、一部の種で確認されている。その際には、体が横分裂によって後部の小さな断片と前部の大きな断片に分かれ、それぞれが失った部分を再生させて2個体になる[1][10]。 フロリダに多産するエダホシムシ属のThemiste lageniformisは、雌だけで単為生殖をすることがある[1][10]。この種では、雄は雌に比べて数が少ない[10]。発生[編集]
海水中で受精した卵は、全割の螺旋卵割を経て胚発生を進める。ごく一部の種は直接発生で、受精卵はゼリー状の物質に覆われたまま発生が進み、小さな蠕虫になって孵化する。しかし大多数は間接発生で、受精卵はトロコフォア幼生となる。トロコフォアは摂食せず、卵黄の栄養を使って発生を進める。トロコフォア幼生から直接に変態して蠕虫になるものもいるが、さらにペラゴスフェラ幼生の段階を経るものもいる。ペラゴスフェラ幼生には卵黄栄養性のものと、プランクトンを摂食するものがいる[1][3]。系統進化[編集]
星口動物は旧口動物であり、そのなかでも環形動物、軟体動物、ユムシ動物などとともに冠輪動物と呼ばれる系統群に含まれる[1]。冠輪動物のなかでは、形態の類似に基づいて軟体動物に近縁と考える意見もあるが、分子系統学の研究は環形動物との類縁を支持するものが多い[11][12]。環形動物に近いというだけでなく、星口動物は系統的に環形動物に含まれるとする結果も得られている[13][14][15]。ストラックらの推定によれば、星口動物は、環形動物のなかではツバサゴカイ類、スイクチムシ類に次いで初期に分岐した系統とされる︵ただし、スイクチムシの位置は不確定︶[14]。形態と分子系統を総合した分析によって、ディノフィリス科の姉妹群と推定した研究もある[16]。 環形動物の体は体節に分かれるが、星口動物に体節はない。星口動物が環形動物に含まれるとする説に基づけば、星口動物が体節を持たないのは祖先的な特徴ではなく、一度体節を進化させた後に失ったのだと考えられる[13][14]。ヤマトサメハダホシムシの神経系の発生過程は環形動物と共通していて、やはり星口動物の祖先は体節を持っていたことを示唆している[17][18]。化石記録[編集]

分類[編集]
ホシムシ類はリンネの﹃自然の体系﹄第12版︵1767年︶に記載されているが、雑多な無脊椎動物とともに蠕虫綱の一員とされていて、独立した分類群にはなっていない。ラマルクやキュビエは、ホシムシ類はナマコに近いと考えていた。1828年にブランヴィルは、ホシムシ類を﹁Sipunculida﹂と名付けたが、このグループには一部の寄生性蠕虫も含まれていた。カトルファージュは、ホシムシとユムシ、エラヒキムシが環形動物と棘皮動物の中間形であると考え、これらの動物をまとめて擬環虫類︵Gephyrea、ギリシャ語で橋を意味するgephyraに由来︶と呼んだ。1959年、ハイマンは星口動物を独立の動物門とした[1]。 ハイマンは星口動物門を綱や目に分けていなかったが、その後の研究により、以下の2綱4目6科に分類されている[1][3]。属以下の分類は各科の記事を参照のこと。各分類群の特徴は西川[5][3]、ブルスカとブルスカ[1]による。 ●スジホシムシ綱 Sipunculidea - 触手は口を囲むように配列する。吻の鉤は輪状に並ばない。 ●スジホシムシ目 Sipunculiformes - 縦筋は分離する。腎管は2本ある。 ●スジホシムシ科 Sipunculidae ●フクロホシムシ目 Golfingiiformes - 縦筋は分離しない。 ●フクロホシムシ科 Golfingiidae - 触手は枝分かれせず、腎管は2本ある。 ●マキガイホシムシ科 Phascolionidae - 触手は枝分かれせず、腎管は1本のみ。 ●エダホシムシ科 Themistidae - 触手は枝分かれし、腎管は2本ある。 ●サメハダホシムシ綱 Phascolosomatidea - 触手は口を囲まず、吻の鉤は輪状に配列する。 ●サメハダホシムシ目 Phascolosomatiformes - 体幹に盾状部を持たない。 ●サメハダホシムシ科 Phascolosomatidae ●タテホシムシ目 Aspidosiphoniformes - 体幹に硬い盾状部を持つ。 ●タテホシムシ科 Aspidosiphonidae 上記の分類体系は、形態による系統推定の結果とほぼ一致する[3]が、分子系統学に基づき、一部見直す必要があるとする研究者もいる[23]。2012年にはやはり分子系統に基づいて科レベルの分類を再編する論文が発表された[24]。それによると、星口動物は以下の6科に分類される。 ●スジホシムシ科 Sipunculidae ●フクロホシムシ科 Golfingiidae ●タテホシムシ科 Aspidosiphonidae ●サメハダホシムシ科 Phascolosomatidae ●Siphonosomatidae ●Antillesomatidae利害[編集]


脚注[編集]
(一)^ abcdefghijklmnopqrstuvwxyzaaabBrusca, Richard C.; Brusca, Gary J. (2003). Invertebrates (2nd ed.). Sinauer Associates, Inc.. pp. 445-451, p.883. ISBN 9780878930975
(二)^ abcdefgh岡田要﹃新日本動物図鑑﹄ 上︵第7版︶、北隆館、1981年、476-478頁。
(三)^ abcdefghijk西川輝昭 著﹁星口動物門﹂、白山義久 編﹃無脊椎動物の多様性と系統︵節足動物を除く︶﹄岩槻邦男・馬渡峻輔︵監修︶、裳華房、2000年、193-195頁。ISBN 4785358289。
(四)^ 白山義久﹁総合的観点からみた無脊椎動物の多様性と系統﹂﹃無脊椎動物の多様性と系統︵節足動物を除く︶﹄、24頁。
(五)^ abc西川輝昭 著﹁星口動物門 Sipuncula﹂、西村三郎︵編著︶ 編﹃原色検索 日本海岸動物図鑑﹄ I、保育社、1992年、299-303頁。ISBN 4586302011。
(六)^ abcdeHoeksema, B.W.; Best, M.B. (1991). “New observations on scleractinian corals from Indonesia:2. Sipunculan-associated species belonging to the genera Heterocyathus and Heteropsammia”. Zoologische Mededelingen 65 (16): 221-245. ISSN 0024-0672.
(七)^ abc本川達雄﹃サンゴとサンゴ礁のはなし﹄中央公論新社︿中公新書﹀、2008年、176-180頁。ISBN 9784121019530。
(八)^ abStolarski, Jarosław; Zibrowius, Helmut; Löser (2001). “Antiquity of the scleractinian-sipunculan symbiosis” (PDF). Acta Palaeontologica Polonica 42 (3): 309-330.
(九)^ Janssen, H. (1990). “Sipunculids from the reef off Mactan Island (Visayas, Central Philippines)”. The Philippine Scientist 27: 11-18.
(十)^ abcdefCutler, Edward B. (1994). The Sipuncula: their systematics, biology, and evolution. Cornell University Press. pp. 298-300, pp.308-310, p.335. ISBN 0801428432
(11)^ Halanych, Kenneth M. (2004). “The new view of animal phylogeny” (PDF). Annual Review of Ecology, Evolution, and Systematics 35: 229-256. doi:10.1146/annurev.ecolsys.35.112202.130124. オリジナルの2008年10月7日時点におけるアーカイブ。.
(12)^ Boore, Jeffrey L.; Staton, Joseph L. (2002). “The mitochondrial genome of the sipunculid Phascolopsis gouldii supports its association with Annelida rather than Mollusca”. Molecular Biology and Evolution 19 (2): 127-137. ISSN 1537-1719. PMID 11801741.
(13)^ abStruck, Torsten H.; Schult, Nancy; Kusen, Tiffany; Hickman, Emily; Bleidorn, Christoph; McHugh, Damhnait; Halanych, Kenneth M. (2007). “Annelid phylogeny and the status of Sipuncula and Echiura”. BMC Evolutionary Biology 7: 57. doi:10.1186/1471-2148-7-57. ISSN 1471-2148.
(14)^ abcStruck, Torsten H.; Paul, Christiane; Hill, Natascha; Hartmann, Stefanie; Hösel, Christoph; Kube, Michael; Lieb, Bernhard; Meyer, Achim et al. (2011). “Phylogenomic analyses unravel annelid evolution”. Nature 471 (7336): 95-98. doi:10.1038/nature09864. ISSN 0028-0836. PMID 21368831.
(15)^ Mwinyi, Adina; Meyer, Achim; Bleidorn, Christoph; Lieb, Bernhard; Bartolomaeus, Thomas; Podsiadlowski, Lars (2009). “Mitochondrial genome sequence and gene order of Sipunculus nudus give additional support for an inclusion of Sipuncula into Annelida”. BMC Genomics 10: 27. doi:10.1186/1471-2164-10-27. PMC 2639372.
(16)^ Zrzavý, Jan; Říha, Pavel; Piálek, Lubomír; Janouškovec, Jan (2009). “Phylogeny of Annelida (Lophotrochozoa): total-evidence analysis of morphology and six genes”. BMC Evolutionary Biology 9: 189. doi:10.1186/1471-2148-9-189. PMID 19660115.
(17)^ Kristof, Alen; Wollesen, Tim; Wanninger, Andreas. “Segmental mode of neural patterning in Sipuncula” (PDF). Current Biology 18 (15): 1129-1132. doi:10.1016/j.cub.2008.06.066.
(18)^ Wanninger, Andreas; Kristof, Alen; Brinkmann, Nora (2009). “Sipunculans and segmentation”. Communicative & Integrative Biology 2 (1): 56-59. doi:10.4161/cib.2.1.7505. PMC 2649304.
(19)^ abcdefHuang, Di-Ying; Chen, Jun-Yuan; Vannier, Jean; Saiz Salinas, J.I. (2004). “Early Cambrian sipunculan worms from southwest China” (PDF). Proceedings of the Royal Society of London. Series B 271 (1549): 1671-1676. doi:10.1098/rspb.2004.2774. ISSN 0962-8452. PMC 1691784.
(20)^ abcdChen, Jun-Yuan (2012). “Evolutionary Scenario of the Early History of the Animal Kingdom: Evidence from Precambrian (Ediacaran) Weng'an and Early Cambrian Maotianshan Biotas, China”. Earth and Life. International Year of Planet Earth. pp. 276-278. ISBN 9789048134281
(21)^ フォーティ, リチャード 著、矢野真千子 訳﹃︿生きた化石﹀生命40億年史﹄筑摩書房、2014年︵原著2011年︶、175頁。ISBN 9784480015884。
(22)^ abMuir, L. A.; Botting, J. P. (2007). “A Lower Carboniferous sipunculan from the Granton Shrimp Bed, Edinburgh”. Scottish Journal of Geology 43 (51-56). doi:10.1144/sjg43010051.
(23)^ Schulze, Anja; Cutler, Edward B.; Giribet, Gonzalo (2005). “Reconstructing the phylogeny of the Sipuncula” (PDF). Hydrobiologia 535-536 (1): 277-296. doi:10.1007/s10750-004-4404-3. ISSN 0018-8158. オリジナルの2010年6月2日時点におけるアーカイブ。.
(24)^ Kawauchi, Gisele Y.; Sharma, Prashant P.; Giribet, Gonzalo (2012). “Sipunculan phylogeny based on six genes, with a new classification and the descriptions of two new families”. Zoologica Scripta 41 (2): 186-210. doi:10.1111/j.1463-6409.2011.00507.x.
(25)^ ab周達生﹃中国食探検 食の文化人類学﹄平凡社、1994年、204頁。ISBN 4582828752。
(26)^ 多紀保彦、奥谷喬司・中村庸夫﹃食材魚貝大百科﹄ 第3巻、近江卓︵監修︶、平凡社、2000年、46頁。ISBN 4582545734。
(27)^ ab辻貴志﹁フィリピン・マクタン島沿岸域におけるサルポとタツナミガイの採捕と利用﹂︵PDF︶﹃日本熱帯生態学会ニューズレター﹄第68号、2007年、6-12頁。
(28)^ 野中健一、石川菜央、宮村春菜﹁人と生き物がつくりだす関係の諸側面 : フィリピン・カオハガン島の事例﹂﹃人文論叢 : 三重大学人文学部文化学科研究紀要﹄第20号、2003年、133-143頁、ISSN 0289-7253、NAID 110000961941。
(29)^ 辻貴志 (2019). “タツナミガイ ︵Dolabella auricularia︶の内臓の採取活動と食用慣行”. 人間文化 45: 37-46.
(30)^ 斉藤英俊、丹羽信彰、河合幸一郎、今林博道﹁西日本における釣り餌として流通される水生動物の現状﹂﹃広島大学総合博物館研究報告﹄第3号、2011年、45-57頁、ISSN 1884-4243、NAID 40019245367。
外部リンク[編集]
- 石田惣 (2005年3月3日). “ホシムシの摂餌行動”. 動物行動の映像データベース. 2011年8月18日閲覧。
- 星口動物の摂餌行動の動画が閲覧できる。
| 典拠管理データベース: 国立図書館 |
|
|---|