本項では1. の意味を解説し、特に断りのない限り、後生動物を指すものとする。
動物を扱う学問を動物学といい、動物の生物学的側面に加え、動物と人とのかかわりが対象とされる[4]。動物の研究史についてはこの「動物学」も参照。
胚が形成される過程で、体軸︵たいじく︶という体の向きが決定がなされ、その向きには前後軸︵頭尾軸︶、背腹軸、左右軸の3つの基本的な軸がある[30]。動物のパターン形成において、体軸の決定など細胞に位置情報を与える機能をもつ物質をモルフォゲンと呼ぶ。
前後軸︵ぜんごじく、antero-posterior axis、頭尾軸、一次軸、吻尾軸︶は動物の体制の基本となる軸で、明瞭な背腹軸のない刺胞動物にも見られ、頭部︵口︶から尾部︵肛門︶を貫いている[32]。前後軸の形成にはほとんどの動物︵例えば、脊椎動物やコオロギ︵節足動物︶やプラナリア︵扁形動物︶から刺胞動物まで︶で Wntリガンド︵細胞外分泌性因子︶が関わっており、尾部側で Wnt、頭部側で Wnt 拮抗因子が発現している[32]。ただし、ショウジョウバエ︵節足動物︶では、初期胚において細胞膜の存在しない合胞体として発生する︵表割︶ため、Wnt のような分泌性因子の濃度勾配ではなくビコイド (bicoid) というホメオドメインを持つ転写因子が蛋白質レベルで頭尾軸に沿って濃度勾配を形成し、形態形成が行われる[32]。また、前後軸に沿った分節の形成にもホメオドメインと呼ばれるDNA結合ドメインを共通に持っている Hox クラスター遺伝子が働いており、胚発生が進むにつれ、遺伝子座の 3'-側から順に前後軸に沿って分節的に発現することで前後軸に沿ったそれぞれの位置に固有な形態が形成される[32]。Hox 遺伝子群は海綿動物をのぞくほぼすべての後生動物が持っている。
背腹軸︵はいふくじく、dorso-ventral axis︶も同様に左右相称動物で認められる動物の体制の基本となる体軸である[32]。扁形動物、節足動物、棘皮動物、脊椎動物など多くの動物で、細胞外に放出される BMP︵骨形成因子︶というリガンドと Chordin などの BMP拮抗因子によってつくられるBMP活性の濃度勾配によって背腹軸が形成される[32]。外胚葉はBMP活性が高いと表皮に、低いと神経に分化するが、19世紀前半から脊椎動物と他の動物では背腹軸に沿った器官配置が反転していることが指摘されており、実際に脊椎動物でBMP が腹側で発現し、背側で Chordin などが発現するのに対し、節足動物︵ショウジョウバエ︶では背側で BMP に相同な分子 (Dpp, Decapentaplegic) が、腹側で BMP拮抗因子︵同、Sog︶が発現していることが分かっている[32]。逆にショウジョウバエにおける腹側を決めるのは dorsal 遺伝子で、細胞性胞胚期において腹側に転写因子ドーサル蛋白質 (Dorsal) が多く分布し、背側への分化を抑制する。胚発生時から背腹軸が決まっている節足動物とは異なり、両生類︵脊椎動物︶では、受精の際に精子の侵入と反対側に灰色三日月環が形成され、そこから原腸陥入が起こって Wnt シグナル伝達系のディシェベルド (Dsh, Dishevelled) が活性化して他の因子を活性化し、反応の下流でオーガナイザーを誘導することで背側となる。
さらに、脊椎動物の神経管の背腹軸は、胚の背腹軸形成の完成後に進行するが、神経管の腹側領域︵フロアプレート︶や脊索で Shh (sonic hedgehog) 蛋白質、Wnt 拮抗因子、BMP拮抗因子が発現し、これらの濃度勾配によって神経管内で下流標的因子の発現活性が活性化または抑制されることで種々の神経細胞が分化する[32]。これらの発現パターンは左右相称動物の中枢神経系で広く保存されている[32]。
左右軸︵さゆうじく︶は動物の3体軸のうち最後に決まる軸で、左右非対称性が生じるメカニズムは進化的に多様である[30]。脊椎動物ではまず胚の中央部︵ノード︶で繊毛の回転により左右対称性が破られ、左側の中胚葉で Nodal および Leftyといったシグナル分子が活性化し、腹腔内で臓器が非対称な形と位置で形成される[30]。それに対し、ショウジョウバエ︵節足動物︶では、細胞の形態のゆがみに起因して消化管が非対称な形態をとる[30]。腹足類︵軟体動物︶では殻の巻く方向が発生初期の卵割様式に依存して Nodal や Pitx2 などの因子の制御により左巻きか右巻きかが変化する[30]。
受精卵が卵割を繰り返し形成される細胞の層を胚葉︵はいよう、germ layer︶と呼ぶ[35]。個体発生の過程では、上皮細胞の層に囲まれ体内と体外の区別がつく胞胚の状態から、原腸陥入によって内胚葉︵ないはいよう、endoderm︶と外胚葉︵がいはいよう、ectoderm︶が形成され、二胚葉性の嚢胚︵原腸胚︶となる[35]。そこからさらに内外両胚葉の何れかから中に細胞が零れ落ち、中胚葉︵ちゅうはいよう、mesoderm︶が形成される[35]。外胚葉由来の中胚葉を外中胚葉︵がいちゅうはいよう、ectomesoderm[注釈7]︶、内胚葉由来の中胚葉を内中胚葉︵ないちゅうはいよう、entomesoderm[注釈8]︶と呼ぶこともある[35]。外中胚葉からなる細胞は全て間充織細胞 (mesenchyme) としてできるが、棘皮動物や箒虫動物など、内中胚葉でも間充織細胞として形成されるものもある。
系統進化の仮説において、多細胞化して細胞同士の密着により体内と外界を隔離するようになった動物が、口と消化管を生じ、内胚葉と外胚葉の区別がなされるようになった二胚葉動物︵ディプロブラスティカ、Diploblastica︶となり、それが更に中胚葉ができて三胚葉動物︵トリプロブラスティカ、Triploblastica︶となったと考えられている。海綿動物以外の動物は︵二次的に喪失したものを除き︶胚葉の分化がみられ、真正後生動物と呼ばれる[38]。刺胞動物および有櫛動物は内中胚葉を持たないため、かつては二胚葉動物と見なされてきたが、内胚葉と外胚葉の間に外中胚葉による間充織細胞を持つため、結合組織に細胞がみられないヒドロ虫類を除き、三胚葉性であるとみなされることが多い︵それぞれの動物門については#現生の動物の系統も参照︶[35]。平板動物も中胚葉を欠くとされるが、前者には上皮の下に細胞がみられる[35][要校閲]。二胚動物および直泳動物にも中胚葉がなく、後生動物ですらない中生動物とされていたが、現在では退化的に単純な体制になったと解釈されている[35]。
 左から無体腔、真体腔、偽体腔の断面の模式図。
左から無体腔、真体腔、偽体腔の断面の模式図。
外胚葉と内胚葉の間隙に中胚葉が筒状の細胞層を形成したものを体腔︵たいこう、coelom︶と呼ぶ[39]。
三胚葉性動物は体腔の構造により、体腔のない無体腔動物︵むたいこうどうぶつ、acoelomates︶、体腔が上皮性の細胞で裏打ちされていない偽体腔動物︵ぎたいこうどうぶつ、pseudocoelomates︶、上皮性の細胞で裏打ちされた体腔をもつ真体腔動物︵しんたいこうどうぶつ、coelomates, eucoelomates︶に大別されてきた[39]。偽体腔は胞胚腔︵原体腔、primary body cavity︶が体腔として残ったもので大きな体腔を作ることができないのに対し、真体腔はしっかりとした大きな体腔を作ることができる。偽体腔動物は従来、袋形動物という一つの動物門に含められていた[39]。また、真体腔はでき方により腸体腔︵ちょうたいこう、enterocoel︶および裂体腔︵れったいこう、schizocoel︶に分けられる[39]。前者は腸体腔嚢と呼ばれる腸管にできる膨らみが括れて切れて形成されるのに対し、後者は中胚葉性の細胞塊の内部に空所が形成される[39]。主に前口動物︵担輪動物 + 脱皮動物︶では裂体腔、後口動物︵新口動物︶では腸体腔となる︵#現生の動物の系統も参照︶。かつて後口動物として扱われていた毛顎動物や腕足動物も腸体腔を持つ。
古くは無体腔動物から偽体腔動物、そして偽体腔動物が真体腔動物に進化してきたと解釈されていたが、ロレンツェン (1985) は間隙生活などで不必要になった真体腔が偽体腔に退化した可能性を示唆しており、さらに分子系統解析の結果でもこれが支持され、無体腔や偽体腔は真体腔が退化的に変化したものである考えがなされている[38][39]。
また、軟体動物、節足動物、尾索動物などでは、血液︵血リンパ︶に満たされた血体腔︵けったいこう、hemocoel︶と呼ばれる腔所︵原体腔︶を持つ。血体腔を持つ動物は開放血管系を持つ。
動物の細胞は、全ての真核生物の細胞に共通した以下の構造を持つ。
●細胞膜‥細胞を包んでいる膜[43]。内部は生体物質を含む水溶液があり代謝の場となっている。リボソーム、細胞質︵原形質︶といった共通の構成要素を持っている。
●DNA‥塩基配列または遺伝暗号 (genetic code) と言うヌクレオチドの塩基部分が並ぶ構造を持ち[44]、遺伝情報の継承と発現を担う。真核細胞のDNAは、一本または複数本の分子から構成される直線状で原核生物よりも多く[45]、染色体と呼ばれる[46]。
●細胞質‥細胞の細胞膜で囲まれた部分である原形質のうち、細胞核以外の領域のこと。真核細胞の細胞質には細胞骨格︵サイトスケルトン︶と呼ばれる微小な管やフィラメント状がつくる網目もしくは束状をした3次元構造[47] がある。これが特に発達した動物の細胞では、細胞骨格が各細胞の形を決定づける。
 トンボの交尾
トンボの交尾
一部の例外を除き動物は何らかの形で有性生殖を行う[54][55]。有性生殖では、減数分裂により一倍体の大小2種類の配偶子が作られる[55]。2つの配偶子が融合する事で新しい個体が生まれるが、この場合小さくて運動性がある配偶子を精子、大きくて運動性を持たない配偶子を卵︵卵子︶といい、配偶子が融合する過程を受精 (fertilization)、受精の結果できあがった細胞を受精卵 (fertilized egg) という[56]。また精子を作る性機能を雄、卵を作る性機能を雌という。雌雄の性機能を別々の個体が担うことを雌雄異体、1つの個体が両方の性機能をもつ場合は雌雄同体であるという。
 脊索動物の初期発生。1: 受精卵、2: 2細胞期、3: 4細胞期、4: 8細胞期、5: 桑実胚期、6: 胞胚期
脊索動物の初期発生。1: 受精卵、2: 2細胞期、3: 4細胞期、4: 8細胞期、5: 桑実胚期、6: 胞胚期
A︵左︶:頭索動物の卵割︵等黄卵︶
B︵中︶:両生類の卵割︵中黄卵︶
C︵右︶:鳥類の卵割︵盤割︶
受精卵や無性生殖におけるなんらかの細胞塊が成体に到達する過程のことを発生'︵はっせい、development︶と呼ぶ。有性生殖では、一倍体である精子と卵︵未受精卵︶が受精する事で、二倍体の受精卵が形成され、発生が開始する。精子由来のミトコンドリアは酵素により分解されるので、ミトコンドリアなどの細胞小器官や母性因子と呼ばれるmRNA、機能タンパク質は卵細胞のみから受精卵に伝わり[61]、子の表現型は母親の影響を受ける母性効果 (materal effect) が現れる。胚発生以前から卵には極性︵軸性、polarity︶があり、卵前核に近い方の極を動物極︵どうぶつきょく、animal pole︶、そうでない極を植物極︵しょくぶつきょく、vegetal pole︶と呼ぶ。前者は幼生の中でも運動や感覚に関する部分、後者は消化器系となり、これらがかつてそれぞれ動物的機能と植物的機能と呼ばれていたためこれらの名がある。
発生が進行すると、胚のそれぞれの部分は特定の組織になるが、その決められた先を予定運命︵よていうんめい、presumptive fate︶と呼ぶ。ある動物において、初期の発生︵2細胞期や4細胞期︶では等しい分化能力︵全能性︶を持ち、すべての組織や器官を形成し得る[38]。ウニの2細胞期の各割球を分けると、それぞれ受精卵と同様に発生が進行する。逆に、4細胞期の環形動物や軟体動物の割球は完全な胚にならない。発生運命が不可逆的に決まることを決定︵けってい、determination︶といい、前者のような状態を﹁未決定である﹂(indeterminate, adj.)、後者のような状態を﹁決定している﹂(determinate, adj.) と表現する。胚発生における発生運命の限定には可逆的に限定された指定 (specification) と不可逆的な決定があり、普通は指定ののちに決定が起こる。Conklin は胚発生の初期において、予定運命の決定が早い段階で起こるものをモザイク卵︵モザイクらん、mosaic egg︶、発生運命が未決定で、各部が影響を及ぼしあいながら順次決まっていくものを調整卵︵ちょうせいらん、regulative egg︶と呼んだ。前者には有櫛動物、紐形動物、線形動物、環形動物、節足動物、軟体動物、尾索動物が、後者には刺胞動物、紐形動物、棘皮動物、腸鰓類︵半索動物︶、脊椎動物などが挙げられる。
受精卵は卵割︵らんかつ、cleavage︶という体細胞分裂を繰り返す事で多細胞からなる胚を形成する。一般的た体細胞分裂とは異なり、卵割の際は核は複製されるが細胞質は卵細胞のものを分割して使うという特徴がある。卵割は分裂溝︵ぶんれつこう、cleavage furrow︶により細胞が2つの割球︵かっきゅう、blastomere︶と呼ばれる細胞に分割されておこる。卵割という用語は受精卵の最初の数回の分割に対して使われる。
卵割様式は卵黄の蓄積部位の影響を受ける。棘皮動物・毛顎動物のように卵黄が等しく分布する等黄卵 (homolecithal egg[注釈9])の場合は、ウニのように等割︵とうかつ、equal cleavage︶を行うか、環形動物や多くの軟体動物のように不等割︵ふとうかつ、unequal cleavage︶となる。これらは卵割面が割球同士を完全に仕切るため全割と呼ばれる。それに対し、端黄卵︵たんおうらん、telolecithal egg︶では分裂溝が卵黄の少ない動物極から現れるため、ハート形分裂︵クラゲ型分裂‥刺胞動物︶の時期を経る。クラゲ型分裂がより極端になると、頭足類︵軟体動物︶のように最初の分裂溝が植物極に達しないまま次の分裂溝が動物極に現れる盤割︵ばんかつ、discoidal cleavage︶を行う。節足動物やイソギンチャク︵のように多量の卵が中央にたまっている心黄卵[注釈10] (centrolecithal egg)では、表割︵ひょうかつ、superficial cleavage︶が行われる。第3分裂︵4細胞期から8細胞期︶では、不等割を行うものでは動物極側のものは小さく、植物極側のものは大きいため、それぞれ小割球︵しょうかっきゅう、micromere︶と大割球︵だいかっきゅう、macromere︶と呼ばれる。
また、卵割では分裂ごとに紡錘体のとる位置や方向が定まっているためそれぞれの分裂方向が一定しており、大きく分けて放射卵割︵ほうしゃらんかつ、radial cleavage︶と螺旋卵割︵らせんらんかつ、spiral cleavage︶の2つの卵割配置 (cleavage pattern) がある。放射卵割では、各分裂の分裂面がその前の分裂に対して直角に起こり、分裂面は卵軸に対して平行か直角に規則正しく起こる。8細胞期以降は不規則な分裂が混ざってくるものが多い。分類群としては、刺胞動物、有櫛動物、箒虫動物、ウニ類︵棘皮動物︶、毛顎動物[75]、腕足動物[75] が挙げられる。螺旋卵割では4細胞期から8細胞期︵第3分裂︶に紡錘体が卵軸に対し45°の角度をなして斜めに位置する[75]。その後の各分裂はだいたい互いに直角に行われるが、初めの分裂面が卵軸に対し傾いているため、以降の分裂面もすべて卵軸に対して角度をなして交わり、螺旋状に並ぶ。分類群としては、扁形動物[75]、環形動物[75]、軟体動物[75] に代表され、紐形動物[75]、内肛動物[75] など少なくとも8つの門が螺旋卵割を行う[75]。なお、環形動物および軟体動物の一部では極体放出および卵割と同期して植物極の細胞質が縊り出され、無核の極葉形成︵きょくようけいせい、polar lobe formation︶が起こる。極葉は一方の割球と合併され、その細胞質は将来の中胚葉となる。8細胞期で大割球から縊り出された4個一組の小割球は第一クオテット︵第一四つ組、1st quartette︶と呼ばれる。また、4細胞期の各細胞からつながる細胞系譜を持つそれぞれの系統をクアドラント︵四分区、quadrant︶と呼ぶ。なお、節足動物などではこのどちらにも当てはまらない[76]。
卵割が進み、細胞が小さくなって胚表面が上皮的に滑らかになると卵割期から胞胚期に移行したとみなされる。この時期の胚は1層の細胞層で囲まれた球形で、胞胚︵ほうはい、(blastula︶と呼ばれる。初期胚の内部には卵割腔が形成されるが、細胞数が増加することで細胞同士が密着結合を形成すると、卵割腔内にNa+やCl-といったイオンが能動輸送され、浸透圧が上昇して内部から水が浸入し胞胚腔液で満たされる大きな胞胚腔 (blastocoel[注釈11]) が形成される[61]。卵割腔︵胞胚腔︶をもつ胞胚を特に中空胞胚 (coeloblastula) と呼び、不等割を行う胚では胞胚の内部は卵黄を含んだ植物極側の大きな細胞で満たされるため中実胞胚 (stereoblastula) と呼ばれる。卵黄量の多い盤割をするものでは細胞は動物極側に偏った胚盤︵はいばん、blatodisc︶を形成し、そのような胞胚を盤胞胚︵ばんほうはい、discoblastula︶と呼ぶ。また表割を行う胞胚では細胞形成は胚の外周でのみ行われるため、囲胞胚︵いほうはい、periblastula︶と呼ばれる。
なお、昆虫や両生類など多くの動物では、卵割期の細胞増殖を急激に行うために通常の細胞分裂で行われる一部の過程︵G1期とG2期の過程︶が省略され早い細胞分裂が続くが[76]、胞胚中期になるとこの省略が終わり、形態形成に必要な転写、細胞の移動や誘導が始まる中期胞胚遷移︵中期胞胚転移、中期胞胚変移︶が起こる[76]。それに対し哺乳類では分裂速度が遅く、2細胞期から既に転写が始まる[76]。
 被いかぶせによる嚢胚形成。
被いかぶせによる嚢胚形成。
1, 4: 外胚葉、2, 5: 内胚葉、3: 胞胚腔、6: 原口
胞胚は内胚葉が外胚葉から分画される嚢胚形成︵原腸胚形成[61]、gastrulation︶を経て嚢胚︵原腸胚[61]、gastrula︶期に至る。嚢胚は内外二重の細胞層からなり、胚葉の区別が現れる。嚢胚を形成する方法は分類群により異なり、最も一般的なものは陥入︵かんにゅう、invagination、または まくれこみ emboly︶である。陥入では植物極側の細胞層が胞胚腔に向かって折れ曲がり、内胚葉となる。内胚葉のつくられた盲管状の部分を原腸︵げんちょう、archenteron︶、その入口を原口︵げんこう、blastopore︶と呼ぶ。この嚢胚形成の方法は棘皮動物などに典型的で、棘皮動物では原腸の両壁には広い胞胚腔が残されているが、箒虫動物では原腸の壁に外肺葉が密着し、胞胚腔を残さない。以降に示す被いかぶせや内展も陥入の変形とみられている。環形動物や軟体動物では被いかぶせ︵おおいかぶせ、epiboly︶という方法で嚢胚形成が行われる。胞胚における動物極側の小割球の分裂が先に進行して、卵黄に富んだ植物極側の大割球を包囲することによって嚢胚ができる。小割球由来の外側の細胞が外胚葉層となり、内側の大割球群が内胚葉となる。被いかぶせでは、胞胚腔はかなり縮小している。また、内胚葉細胞塊ははじめ原腸を形成しないため、外胚葉に覆われていない部分を原口と呼んでいるが、発生の進行に伴って原腸を形成し、原口と連絡する。この場合、原口から落ち込んだ外胚葉の細胞層を、口陥︵こうかん、stomodaeum︶と呼ぶ。盤胞胚を形成する頭足類では、胚盤葉の一端がその下に折れ込んで前方に延長する内展︵ないてん、involution︶によって内胚葉が形成される。
もう一方の嚢胚形成の方法は葉裂法︵ようれつほう、delamination︶と呼ばれ、主に刺胞動物にみられる。狭義の葉裂法はカラカサクラゲ類 Geryoniidae にのみ見られ、中空胞胚において外壁を作る細胞が一様に胞胚腔に向かって分裂すると、胞胚腔内に出た細胞は規則正しく配列して内胚葉の嚢を作る。ヒドラなどが行う方法は多極法︵たきょくほう、multiopolar proliferation︶と呼ばれ、胞胚法を形成している細胞が各所で胞胚腔内にすべり落ち、それが内胚葉の嚢を形成する。それに対し、ウミコップ属 Clytia では植物極のみから細胞がすべり落ちるため、単極法︵たんきょくほう、uniopolar proliferation︶と呼ばれ、多極法と併せて極増法︵きょくぞうほう、polarization︶と呼ばれる。葉裂法を行う嚢胚の多くは中実嚢胚︵ちゅうじつのうはい、stereogastrula︶で、発生が進行するまで原腸も原口も持たない。
左右相称動物では、内胚葉および外胚葉とは別に、体腔と関連して中胚葉の形成が起こる。刺胞動物や有櫛動物では外肺葉から細胞が零れ落ち、外中胚葉性の間充織細胞を作る。棘皮動物や箒虫動物など、内中胚葉でも間充織細胞として形成されるものはあるが、内中胚葉は普通表皮の形をとる。
螺旋動物では、まず第二クオテットまたは第三クオテットから外中胚葉性の間充織細胞が形成される。その後、D四分区の 4d細胞︵中胚葉帯端細胞、mesoblastic teloblast︶から内胚葉由来の中胚葉が生まれる。第四クオテットの他の細胞︵4a, 4b, 4c︶は内胚葉となる。かつては 4d細胞の系統にある子孫細胞は全て中胚葉になると考えられていたが、内胚葉も含んでいる。4d細胞は胞胚腔内に落ちると左右に分裂し、胚の分化に伴い肛門になる部分の左右前方に位置しながら前方に細胞を送り、中胚葉帯︵mesoderm band︶を作る。これを﹁端細胞による中胚葉形成法 telobblstic method﹂と呼ぶ。環形動物などでは、この中胚葉帯内に体腔が形成され、これが裂体腔と呼ばれる。
節足動物でも、中胚葉は1対の細胞帯として出現する。しかし螺旋動物のように特定の細胞ではなく、原口の周囲の細胞群に由来している。
腸体腔をもつ後口動物および毛顎動物、腕足動物などでは、原腸壁の一部が胞胚腔に向かって膨出 (evagination) し、そこから分離して胞胚腔内で独立した体腔嚢︵たいこうのう、coelomic vesicle︶を形成する。こうしてできた体腔は腸体腔であり、それを囲む壁が中胚葉である。脊椎動物においては、両生類︵無羊膜類︶では中胚葉の形成と原腸の形成が同時に起こるが、羊膜類︵鳥類や哺乳類︶では、中胚葉の形成が先に行われ、その後卵黄嚢と連続する内胚葉の一部が中胚葉に包み込まれるようにしてくびれ、原腸の形成が行われる[61]。
脊椎動物などでは、組織や器官を形成するため、胚細胞が特定の機能を持った細胞に変化する︵細胞分化︶。この際、基本的な細胞機能の維持に必要な遺伝子︵ハウスキーピング遺伝子︶の機能は残しつつ、特定の機能に必要な遺伝子を新たに発現し、逆に分化後には不必要になる遺伝子をDNAメチル化により不活性化する。
脊椎動物などでは原腸胚期の後、神経管が形成される神経胚期へと進む。例えばニワトリでは、外胚葉に神経板という領域ができ、それが胚の内側に丸まる事で神経管ができ、さらに直下に脊索が形成される。神経管の前方には前脳、中脳、後脳という3つの膨らみが形成され、これらが将来脳になる。脊索の両側の沿軸中胚葉から体節が形成され、体節と隣接した外側の中間中胚葉からは腎節が形成される。体節はやがて皮節、筋節、硬節に分かれ、これらはそれぞれ皮膚の真皮層、骨格筋、椎骨などが形成され、腎節からは腎臓や生殖腺が形成される。中間中胚葉のさらに外側には予定心臓中胚葉という、将来心臓関連の組織になる部分があり、これは壁側中胚葉と臓側中胚葉に転移する。前者からは体腔を覆う胸膜や腹膜が形成され、後者からは心筋、平滑筋、血管、血球などが形成される。心臓は生命の維持に不可欠なので、発生の早い段階で中胚葉から形成される。なお、予定心臓中胚葉は中胚葉の正中線を隔てた両側に2つ存在するが、これら2つは移動して胚の前方で合流して心臓を形成する。脊椎動物では外胚葉と中胚葉の相互作用で四肢が形成される。ヒトの手足は水鳥と違い、指の間に水かきがないが、これはアポトーシスの作用で水かき部分の細胞を﹁自殺﹂させている為である。
動物の起源については、単細胞生物の襟鞭毛虫が集まって多細胞化する事で海綿動物のような動物になっていったと考えられる。これをガストレア説︵群体繊毛虫仮説︶と呼ぶ。ヘッケルは動物の初期発生に基づき、襟鞭毛虫のような原生動物から、胞胚に相当する1層の細胞層を持つ中空の祖先型動物ブラステア (Blastea) が生じ、次に嚢胚に相当する二重の細胞層からなる袋状のガストレア︵腸祖動物、Gastraea︶が生じたと想定した[89]。
なお従来は、上述した襟鞭毛虫類から進化したとするヘッケルの説と繊毛虫類から進化したとするハッジの説︵多核体繊毛虫仮説、合胞体繊毛虫仮説︶が対立していたが、分子遺伝学の成果によれば、18SrDNAに基づいた解析などにより、動物は襟鞭毛虫類を姉妹群に持つ単系統な群であることが示されており、ヘッケルの説が有力とされている[89]。ハッジの説は生態学的な視野のもと、多核繊毛虫から無腸動物のような原始的な左右相称動物が生じたと考え、後生動物の起源を左右相称動物に求めた[89]。
この多細胞化が起こった仮説として、現在までに様々なものが提案されてきた。複雑な多細胞生物の出現は、生物圏の酸化が進むまで妨げられたという説が広く受け入れられてきた。ほかにも動物が多様化するきっかけとしてとして、クライオジェニアンやエディアカラ期の全球凍結の環境的制約から後生動物の祖先が解放されたこと、宇宙放射線の影響、極移動、大陸の分断、硫化水素の毒性、塩分、微量金属の栄養塩の不足、海に栄養塩をもたらす大陸風化の周期、地球温暖化、または活発になった捕食者と捕食者の軍拡競争などが考えられるが、必ずしも相互に排他的なものではない。なおこれらの仮説は、多少なりとも、後生動物の多様化との因果関係につながるが、結局推定される時間的な一致に依存しており、地球規模の海の大酸化は後生動物が進化した原因ではなく、後生動物の出現による結果であると主張されている。
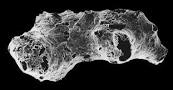 オタヴィアの化石
オタヴィアの化石
 カンブリア紀の生物アノマロカリスの復元図
カンブリア紀の生物アノマロカリスの復元図
 トリケラトプスの骨格化石
トリケラトプスの骨格化石
新生代は哺乳類が優勢になり、鳥類、昆虫類、真骨魚類も適応放散し、現在と同様の動物相が形成された。新生代の後半にあたる第四紀には人類も出現した。
各動物門に含まれる代表的な動物の例(和名は総称、詳細は「種名」を参照)
- 海綿動物門 Porifera Grant, 1836
- 有櫛動物門 Ctenophora Eschscholtz, 1829[注釈 14]
- 刺胞動物門 Cnidaria Verrill, 1865[注釈 14]
- 平板動物門 Placozoa K.G. Grell, 1971(板形動物)
- 珍無腸動物門 Xenacoelomorpha Philippe et al., 2011[注釈 15]
- 棘皮動物門 Echinodermata Leuckart, 1854
- 半索動物門 Hemichordata Bateson, 1885
- 頭索動物門 Cephalochordata Lankester, 1877[注釈 16]
- 尾索動物門 Urochordata Lankester, 1877[注釈 16]
- 脊椎動物門 Vertebrata J-B. Lamarck, 1801(有頭動物 Craniata Lankester, 1877)[注釈 16]
- 毛顎動物門 Chaetognatha Leuckart, 1854
- 胴甲動物門 Loricifera Kristensen, 1983
- 動吻動物門 Kinorhyncha Reinhard, 1887
- 鰓曳動物門 Priapulida Théel, 1906
- 線形動物門 Nematoda Diesing, 1861(Nemata Cobb, 1919)
- 類線形動物門 Nematomorpha Vejedovsky, 1886(Gordiacea von Siebold, 1843)
- 緩歩動物門 Tardigrada Spallanzani, 1777
- 節足動物門 Arthropoda Siebold & Stannius, 1845
- 有爪動物門 Onychophora Grube, 1853
- 直泳動物門 Orthonectida Giard, 1877[注釈 17]
- 二胚動物門 Dicyemida van Beneden, 1876(菱形動物 Rhombozoa van Beneden, 1882)[注釈 17]
- 有輪動物門 Cycliophora Funch & Kristensen, 1995
- 顎口動物門 Gnathostomulida Ax, 1956
- 微顎動物門 Micrognathozoa Kristensen & Funch, 2000
- 輪形動物門 Rotifera Cuvier, 1798[注釈 18]
- 腹毛動物門 Gastrotricha Metschnikoff, 1864
- 扁形動物門 Platyhelminthes Hyman, 1951(Plathelminthes Schneider, 1873)
- 苔虫動物門 Bryozoa (外肛動物 Ectoprocta Nitche, 1870)
- 内肛動物門 Entoprocta Nitche, 1869(曲形動物 Kamptozoa Cori, 1921)
- 箒虫動物門 Phoronida Hatschek, 1888
- 腕足動物門 Brachiopoda A.M.C. Duméril, 1806
- 紐形動物門 Nemertea Quatrefages, 1846(Rhynchocoela Schultze, 1851)
- 軟体動物門 Mollusca Cuvier, 1797
- 環形動物門 Annelida J-B. Lamarck, 1809[注釈 19]
1990年代以前は左右相称動物は原腸が口になるか否かで前口動物、後口動物に分類され、さらに体腔が無体腔、偽体腔、真体腔のいずれであるかにより分類されていた。しかし1990年代の18S rRNA遺伝子の解析により、体腔の違いは進化とは関係ない事が判明し、上述の意味での後口動物は単系統でない事が示されたので、いくつかの動物門を新口動物から外し(後述)、前口動物に移した。このような変更を施した後の前口動物が単系統であることが支持されている[146][156][157]。
下記は主に ギリベ (2016)の系統仮説に基づく系統樹に、ラーマーら (2019) による分子系統解析の結果を加えて、動物界の系統樹を門レベルまで描いたものである[11][158][159][注釈20]。ただし、2018年現在、分子系統解析が進展中ということもあり、完全に合意がなされたものではない。本項はこの系統樹に基づき以下の小節にて解説を行う。
海綿動物門、平板動物門、刺胞動物門、有櫛動物門の4つは左右相称動物に含まれない動物門で、体の左右相称性がなく、これらをまとめて便宜的に﹁前左右相称動物﹂と呼ぶこともある。分子系統解析から、このうち海綿動物か有櫛動物の何れかが後生動物で最も系統の基部に位置すると考えられている[11][159]。しかし、海綿動物が系統の最も基部に位置するか[159][163][164][165][166]、有櫛動物が系統の最も基部に位置するか[167][168][169][170] は分子系統解析においてもデータが分かれている。
現在の多様性は単純なものから複雑なものに進化してきたとする考え方のもと、かつては最も単純な平板動物から、細胞の種類がより多い海綿動物、そして神経を持つ刺胞動物、最後に神経系に加え筋系ももつ有櫛動物が進化してきたと考えられた[11][158]。ただし、襟鞭毛虫との類似から海綿動物のほうがより原始的な姿に近いとする考えもあった[158]。この進化的な仮説は形態に基づく分岐学的解析においても一時は支持された[158]。しかし、分子系統学が導入された初期にはもう平板動物は二次的に退化したより派生的なグループであることが明らかになり、有櫛動物は刺胞動物より系統の基部に位置することが明らかになった[158][171]。それだけでなく、有櫛動物はほかのすべての後生動物よりも基部に分岐したとする結果が得られた[167][168]。海綿動物は相称性や胚葉がなく体制が単純であるため、最も初期に分岐した後生動物として直感的に受け入れられやすいのに対し、有櫛動物は放射相称、神経系と筋系をもつため、有櫛動物より後に海綿動物が分岐したと考えると筋系や神経系が有櫛動物と Parahoxozoa︵有櫛動物と海綿動物以外の後生動物︶で2回独立に獲得したと考えるか、海綿動物でどちらも1回完全に喪失したと考えなければならないため、大いに議論を呼んだ[11]。系統誤差の影響を軽減することで、再び海綿動物が最も初期に分岐したと考えられる結果が得られている[159][166]。
海綿動物 Porifera は相称性がなく胚葉がないなど最も単純なボディプランを持つ。海綿動物の細胞は分化するものの、組織を形成することはなく[172]、複雑な器官をもたない[173]。そういったことから海綿動物は側生動物 Parazoa Sollas, 1884 と呼ばれることもある。
刺胞動物と有櫛動物の体は放射相称性を持ち、唯一の腔所である胃腔の開口は口と肛門を兼ねる。これらの動物門の細胞は組織に分化しているものの、器官を形成していない[175]。中胚葉が形成されない二胚葉性の動物であるとされるが、細胞性である間充織を中胚葉とみなし、ヒドロ虫綱以外の刺胞動物と全ての有櫛動物を三胚葉性とみなす事も多い[176]。
刺胞動物は触手に物理的または化学的刺激により毒を含む刺糸を発射する刺胞と呼ばれる細胞器官を持つ。漂泳性︵クラゲ型︶と付着性︵ポリプ型︶という生活様式の異なる2つの型を持ち雌雄異体である。かつては単細胞生物とも考えられていた寄生性のミクソゾアは分子系統解析により刺胞動物に内包されている[176]。
それに対し有櫛動物は1個の細胞が変形してできた膠胞を持ち、中胚葉性の真の筋肉細胞を持つほか、全てクラゲ型であり、二放射相称で雌雄同体である[178]。
平板動物は神経細胞も筋肉細胞も持たず、体細胞は6種類しかなく器官や前後左右軸をもたない、自由生活を行う動物として最も単純な体制を持つ[176]。しかし2008年にセンモウヒラムシ Trichoplax adherens のゲノム解読がなされ、シグナル伝達系、神経やシナプス、細胞結合などに関する多くの遺伝子の存在が報告された[176]。
4つの門を除いた全ての動物門が左右相称動物である。左右相称動物は完全な三胚葉性で、体が左右相称である。外見上は左右対称であるが、内部の臓器は限られた空間の中に各臓器を互いの連結を保ちながら機能的に配置するために、位置や形が左右非対称となっている[30]。
左右相称動物は口と肛門、およびこれらをつなぐ消化管をもち、体内に体腔ないし偽体腔︵線形動物、輪形動物など︶を持つ。左右相称動物のボディプランは、前方︵運動のとき体の進む方向︶と後方の区別、腹側と背側の区別がある傾向があり、したがって左側と右側の区別も可能である[180][181]。運動のとき体の前方へと進むので、進行方向にあるものを識別する感覚器や餌を食べる口が前方に集まる傾向にある︵頭化という︶。多くの左右相称動物は環状筋と縦走筋のペアを持つので[181]、ミミズのような体が柔らかい動物では流体静力学的骨格︵水力学的骨格、流体包骨格、hydrostatic skeleton︶の蠕動により動く事ができる[182]。また多くの左右相称動物には繊毛で泳ぐことができる幼生の時期がある。
以上の特徴は例外も多い。例えば棘皮動物の成体は︵幼生とは違い︶放射相称であるし、寄生虫の中には極端に単純化された体の構造をもつものも多い[180][181]。
 アワフキムシの脱皮
アワフキムシの脱皮
螺旋動物の系統関係
| ラーマーら (2019) に基づく分子系統樹の例[75][159] |
Marlétaz et al. (2019) に基づく分子系統樹の例[75]
|
|
|
|
 ニシキウズガイ属(軟体動物腹足類)の胚の螺旋卵割
ニシキウズガイ属(軟体動物腹足類)の胚の螺旋卵割
 受精から9時間の海洋性の腹足類 Haliotis asinina のトロコフォア
受精から9時間の海洋性の腹足類 Haliotis asinina のトロコフォア
冠輪動物に属する軟体動物門は節足動物門に次いで既知種の大きい門で、骨格を持たず、体節がない軟体からなる。体腔は真体腔であるが退化的で、体内の腔所は組織の間隙を血液が流れるだけの血体腔である。一般的には体は頭部、内臓塊、足からなり、外套膜が内臓塊を覆っている。外套膜が分泌した石灰質の貝殻を持つ事が多い。卵割は普通全割の螺旋卵割であるが、頭足類では胚盤をもつ盤割となる。
軟体動物の分類は系統解析により一部修正が施され2018年現在は体全体を覆う大きな殻がある有殻類と石灰質の棘を持つ有棘類に大きく分かれるという仮説が有力視されている[230]。
軟体動物の綱は以下のように分類される[230]‥
有殻類は綱レベルの単系統性は多くの場合保証されているが、各綱の系統関係は2018年現在一致を見ていない[230]。
分子系統解析に基づく系統樹は次の通りである[231][注釈 31]:
|
|
| 二胚動物と直泳動物を吸啜動物の姉妹群とする分子系統樹の例[160]
|
分子系統解析から、かつて中生動物とされていた二胚動物および直泳動物はともに螺旋動物に属することが支持されている。ただし、その中でも、二胚動物と直泳動物は姉妹群﹁中生動物﹂となり、さらにそれが吸啜動物と姉妹群をなすという結果もあれば[160]、直泳動物は環形動物に内包され、環形動物の極端に退化した形と考えられることもあり[161]、まだ決着はついていない。
 前口動物︵上図、Protostomes︶と後口動物︵下図、Deuterostomes︶の発生。
前口動物︵上図、Protostomes︶と後口動物︵下図、Deuterostomes︶の発生。
8細胞期 (eight-cell stage) では前者は螺旋卵割 (spiral cleavage)、後者は放射卵割 (radial cleavage) を行う。原腸陥入 (gastrulation)においても体腔 (coelum) のできる位置が異なることが多く、前者では基本的に裂体腔で後者では基本的に腸体腔である[注釈32]。また、名の由来の通り前者では原口 (blastopore) が口 (mouth) となるのに対し、後者では原口が肛門 (anus) となる。
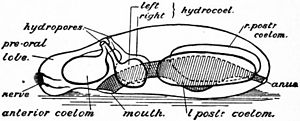 ディプリュールラ幼生。トロコフォア幼生と対置される。
ディプリュールラ幼生。トロコフォア幼生と対置される。
後口動物︵新口動物︶は棘皮動物門、半索動物門、脊索動物を含み、新口動物とも呼ばれる。ヘッケルは新口動物の共通祖先から脊索動物が進化した過程を論じた際、棘皮動物の幼生[注釈33] と半索動物のトルナリア幼生が共有する形質を合わせて、それらの祖先型として、ディプリュールラ幼生 (Dipleurula) という仮想的な幼生を考えた。ディプリュールラ幼生はトロコフォア幼生と同様に口から肛門に至る消化管、頂器官に感覚器としての長い繊毛、口を中心とした繊毛帯︵または繊毛環︶、体後端部の端部繊毛帯を持つが、ディプリュールラ幼生では3部性の体腔︵原体腔・中脳腔・後脳腔︶を持つことおよび繊毛帯の走り方が異なる[235][236]。
2018年現在、棘皮動物と半索動物が姉妹群をなすという説が大勢を締めており[11]、これら2つをあわせて水腔動物 Coelomopora という[11]。
後口動物は胚発生において陥入によってできた原口が口になる前口動物に対し、原口が口にならず新たに口が開く動物であり、かつては現在後口動物とされる棘皮動物、半索動物、脊索動物だけでなく、触手冠動物としてまとめられる箒虫動物、苔虫動物︵外肛動物︶、腕足動物、そして毛顎動物を含んでいた[238]。これはブルスカとブルスカ (1990)、メルグリッチとシュラム (1991)などによる形態形質に基づく系統解析でも、原口に由来しない口を持つだけでなく、原腸由来の中胚葉を持つことや腸体腔を持つことなどの形質からも支持されていた[238]。ほかにも、放射卵割を行うなど[75]、後口動物としての性質を多く持っている。しかし分子系統解析の進展により、触手冠動物および毛顎動物は前口動物に属すると考えられるようになった[75][221]。この変更以降も﹁後口動物﹂という系統群名を用いるが[239][240][241][242][242][243][244][245] 、毛顎動物や腕足動物のような原口が口にならない動物も前口動物に含まれ[75]、単純に原口の有無が系統を反映しているわけではない。
水腔動物 Coelomopora︵歩帯動物 Ambulacraria︶は幼生の形態、三体腔性、軸器官などの形質を共有する[11]。
棘皮動物は、成体が五放射相称、三胚葉性で、内胚葉由来の中胚葉︵内中胚葉︶を持つ。腸体腔性の体腔で、体腔に由来する水管系と呼ばれる独自の構造をもつ[247]。神経系は中枢神経を持たず、神経環と放射神経からなるが、ウミユリ綱では神経節を持つ。ウミユリ綱、ヒトデ綱、クモヒトデ綱、ナマコ綱、ウニ綱からなり、分子系統解析によりこれらのうちウミユリ綱が最も祖先的だと考えられている[247]。ウニ綱のうちタコノマクラ類やブンブク類では五放射相称が歪み左右相称性を示す[247]。
現生の半索動物はギボシムシ綱︵腸鰓綱︶とフサカツギ綱︵翼鰓綱︶からなり、化石ではフデイシ綱が置かれる[249][250]。どちらも体は前体・中体・後体の3つの部分に分かれるという共通した形質を持ち、前者では吻・襟・体幹と呼ばれ、後者では頭盤・頸・体幹と呼ばれる。ギボシムシ綱では腸体腔と裂体腔をもつとされるが、体腔形成には不明な点も多い。ギボシムシ綱は側系統で、ギボシムシ綱のハリマニア科がフサカツギ綱と姉妹群をなし、フサカツギ綱はギボシムシ綱から小型化によって体が二次的に単純化したと考えられる。半索動物は脊索動物と同様に鰓裂を持つ[250]。かつては口盲管という器官が脊索の一種と考えられたこともあったが、口盲管と脊索との関係を支持する発生遺伝学的研究結果はなく、現在では脊索を持たないとされる[250]。
脊索動物 Chordata は頭索動物・尾索動物︵被嚢動物︶・脊椎動物を含むクレードで、一生のうち少なくとも一時期に鰓裂・脊索およびその背側に背側神経管を持つという形質を共有する[250]。脊索は膨らませた細長い風船に喩えられる中軸器官で、脊索鞘という繊維質の頑強な膜に脊索細胞が包まれている[250]。頭索動物および尾索動物がもつ内柱は脊椎動物における甲状腺と相同で、甲状腺は内柱の変化したものと考えられている。発生はさまざまであるが発生の一時期には肛門の後方に筋肉により運動する尾状部分があり、オタマジャクシ型幼生︵tadpole larva︶を経る。
脊索動物は脊索と背側神経管という共通する二つの特徴をもつことから1つの門に置かれ、その中の3群は亜門に置かれてきたが、佐藤矩行・西川輝昭 (2014)により、分子系統学的解析および3群がそれぞれ特徴的な形質を持つことに基づいて脊索動物をより高次の上門に置き、3群を門に格上げする考えが提唱された[250][255][256]。
以下の3つに分類される[11][250]‥
伝統的に諸民族で、生物は植物と動物に大別されてきた。古代ギリシアのアリストテレスは﹃動物誌 Περὶ Τὰ Ζῷα Ἱστορίαι﹄などの著作において動物と植物の中間的存在を認めつつこの区分を採用し、感覚と運動の能力は動物にだけ見られるとし、霊魂の質的差異によって理論的に説明しようとした。さらに動物を赤い血を持つ有血動物︵ἐναίμος、現代の﹁脊椎動物﹂に相当︶とそうでない無血動物︵ἀναίμος、現代の﹁無脊椎動物﹂に相当︶に二分し、発生様式と足の数を主要な基準として体系的に細分した[260]。アリストテレスはリンネ式階層分類とは異なり、全ての上位分類に﹁類 γένος﹂を用い[260]、有血動物を人類・胎生四足類・卵生四足類・鳥類・魚類に、無血動物を軟体類︵μαλάκια、現在の頭足類︶・軟殻類︵軟甲類、μαλακόστρακα、現在の軟甲類 Malacostraca に相当︶・有節類︵ἔντομα、現在の節足動物から甲殻類を除いた概念︶・殻皮類︵ὀστρακόδερμα、現代の貝類に加え、ウニ類、ホヤ類を含む︶に分けた[260]。
| 『自然の体系 初版』(1735) |
『自然の体系 第10版』(1758)
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 粘液胞子虫の一種 Myxobolus spinacurvatura︵ミクソゾア動物︶
粘液胞子虫の一種 Myxobolus spinacurvatura︵ミクソゾア動物︶
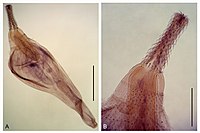 古鉤頭虫綱の一種 Corynosoma cetaceum︵鉤頭動物︶
古鉤頭虫綱の一種 Corynosoma cetaceum︵鉤頭動物︶
 サツマハオリムシ Lamellibrachia satsuma︵有鬚動物‥ハオリムシ動門︶
サツマハオリムシ Lamellibrachia satsuma︵有鬚動物‥ハオリムシ動門︶
 イヌシタムシ Linguatula serrata︵舌形動物︶
研究の進行、特に分子系統解析の台頭により解体または他の動物門の下位に吸収された動物門も多く存在する。詳細は各項を参照。
イヌシタムシ Linguatula serrata︵舌形動物︶
研究の進行、特に分子系統解析の台頭により解体または他の動物門の下位に吸収された動物門も多く存在する。詳細は各項を参照。
腔腸動物門 Coelenterata Hatschek, 1888
現在は刺胞動物門および有櫛動物門に分割されている。かつては胃水管系を腔腸 (coelenteron) と呼び、腔腸動物としてまとめられていた[178]。また放射相称動物 Radiata と呼ばれることもあった。有櫛動物は、細胞器官である刺胞の代わりに1個の細胞が変形してできた膠胞を持つことや、上皮細胞の各細胞が2本以上の繊毛を備える多繊毛性であること、中胚葉性の真の筋肉細胞を持つこと、卵割は決定性卵割であること、複数の感覚器が放射相称的に配置される刺胞動物とは異なり1個のみを反口側に持つことなど、刺胞動物と大きく異なっており、しかも分子系統解析により腔腸動物が単系統とならないことがわかったので両者は別の門として分けられている[178]。
ミクソゾア門 Myxozoa Grassé, 1970
原生動物の一群として扱われることもあったが、極糸が入った極嚢という構造が刺胞に似ており、分子系統解析の結果、現在では刺胞動物に含められる[176][263]。後生動物特有の細胞間接着構造や動物のみに存在するHox 型ホメオティック遺伝子を持ち、寄生性の獲得により二次的に退化した体制となったと考えられている[263]。
中生動物門 Mesozoa van Beneden, 1876
現在は二胚動物門および直泳動物門に分割されている。Édouard van Beneden (1876) により原生動物と後生動物の中間に位置をする動物群として、ニハイチュウ︵二胚動物︶のみを含む群として設立され、のちに van Beneden (1882) にチョクエイチュウ︵直泳動物︶がこれに含められた[264]。その後様々な生物が含められたがその正体が渦鞭毛藻やミクソゾアであることがわかり、除かれた[264]。Kozloff (1990) は、あるステージのニハイチュウ類はチョクエイチュウ類のそれに表面的には似ているが、それ以外の点においては明確に異なっているとして、これらを独立の門に置いた。
一胚葉動物門 Monoblastozoa R. Blackwelder, 1963
1982年にアルゼンチンの岩塩から発見された1層の体皮細胞からなる生物であるが、存在が疑問視されている。
袋形動物門 Aschelminthes taxon inquirendum
偽体腔をもつ動物をまとめた﹁ごみ箱分類群 wastebasket taxon﹂で、現在は輪形動物・鉤頭動物・腹毛動物・線形動物・類線形動物・動吻動物・胴甲動物・鰓曳動物・内肛動物に分割されている[39]。鉤頭動物・線形動物・類線形動物は円形動物としてまとめられたこともあった。
前肛動物門 Prosopygii Lang, 1888
箒虫動物、苔虫動物、腕足動物、ほかにも星口動物およびフサカツギ類などはかつてまとめて前肛動物と呼ばれ1門に置かれていた[267][268]。箒虫動物・苔虫動物・腕足動物の3分類群は現在でも触手冠動物として門より高次の分類群をなすことがある[11]。
鉤頭動物門 Acanthocephala Kohlreuther, 1771
現在は輪形動物に内包され、かつての狭義の輪形動物は側系統となる[153]。狭義の輪形動物および鉤頭動物を門として残し、現在の広義の輪形動物を共皮類︵多核皮動物︶ Syndermata とすることもある[153]。
有鬚動物門 Pogonophora Johansson, 1937
現在は環形動物門に内包されている[269]。狭義の有鬚動物︵ヒゲムシ︶と下記のハオリムシは体後端の体節構造および成体での消化管の喪失などの共有派生形質をもち、まとめて有鬚動物とする考えが主流であった[269]。溝副触手綱 Canalipalpata ケヤリ目 Sabellida に含まれる1科、シボグリヌム科 Siboglinidae となっている。
ハオリムシ動物門 Vestimentifera Webb, 1969
現在は環形動物門に内包されている[269]。もともと上記の有鬚動物に含められていたが、ジョーンズ (1985) は体腔の構造の違いを重視し、独立した門に置いた[269]。しかし、当時よりSouthward (1988) のように反対意見も多く、上記のような共有派生形質を持つことから以降も有鬚動物とされることが多かった[269]。現在は上記のシボグリヌム科に含められる。
星口動物門 Sipuncula Rafinesque, 1814
現在は環形動物門に内包されている。分子系統解析によりフサゴカイ目と姉妹群をなすことが分かった。
ユムシ動物門 Echiura Newby, 1940
現在は環形動物門に内包されている。分子系統解析によりイトゴカイ目に内包されることが分かった。
舌形動物門 Pentastomida Diesing, 1836
現在は節足動物門に内包されている。魚類の外部寄生虫である鰓尾類と近縁であることがわかり、21世紀以降はウオヤドリエビ綱の中の1亜綱、舌虫亜綱 Pentastomida Diesing, 1836 とされる[209]。
単肢動物門 Uniramia[273]
現在は節足動物門に内包されている。昆虫類および多足類を共通の性質を持つとして合わせ、鋏角類や甲殻類とともに独立した門とされることもあった[273]。しかし21世紀以降、昆虫は甲殻類と単系統群の汎甲殻類をなすことが明らかになっており[207]、もはや用いられない。
1960年以降に提唱され、現在も用いられている動物門を挙げる。詳細は各項を参照。
平板動物門 Placozoa Grell, 1971
1883年にオーストラリアの水族館で発見されたが、採集方法が確立し詳細な形態観察できるまで存在が認められなかった[269]。1971年に平板動物門が設立された。
顎口動物門 Gnathostomulida Ax, 1956
アックス (1956) によって発見され扁形動物の1目として記載されたが、リードゥル (1969) により独立の動物門に移された[153][269]。
胴甲動物門 Loricifera Kristensen, 1983
クリステンセン (1983) により記載された[269]。
有輪動物門 Cycliophora Funch & Kristensen, 1995
Funch & Kristensen (1995) により記載された[269]。
微顎動物門 Micrognathozoa Kristensen & Funch, 2000
2000年にグリーンランドの湧水から発見され、担顎動物門の一綱として記載された。
珍無腸動物門 Xenacoelomorpha Philippe et al., 2011
無腸類と皮中神経類を含む無腸動物とチンウズムシの仲間を合わせたクレードである[145]。
人間が野生動物︵原種 original breed︶から遺伝的に改良し、繁殖させて人間の生活に利用する動物を家畜︵かちく、domestic animal︶という[275]。これには哺乳類以外の鳥類・爬虫類や昆虫も含まれるが、特に鳥類を家禽︵かきん、poultry[277], fowl︶として区別することもある[275]。また、広義の家畜は農用動物、愛玩動物、実験動物に大別され、このうちの農用動物のみを指して家畜 (farm animal, livestock) と呼ぶこともある[275]。
愛玩動物 (pet animal) とは、一般に家庭などで愛玩のために飼育されている動物で、特に愛玩飼育を目的として改良・繁殖が行われてきた動物をいう[283]。
展示動物とは、動物園で展示されている動物のように展示を目的として飼育されている動物をいう[283]。
記事冒頭の通り、動物界︵後生動物︶を﹁動物﹂として扱うことが一般的であるが、﹁動物﹂の語は学術的な場面でもほかの語義を持つことがある。
原生動物︵protozoans︶
捕食や移動など、動物的な特徴を持った単細胞や群体性真核生物︵非単系統群︶に対する慣用名。二界説の時代に動物界における原生動物門︵または原生動物亜界 ︶Protozoa とされ、鞭毛虫類、肉質虫類、胞子虫類、繊毛虫類に細分されていた。
動物プランクトン︵zooplankton︶
プランクトン︵浮遊生物︶のうち、鞭毛などにより運動性と持つもので、原生動物、節足動物︵橈脚類・鰓脚類︶、輪形動物を主とする。
動物性機能︵animal function︶
生体の持つ機能のうち、運動・感覚・神経相関の3つを指し、この働きに携わる器官を動物性器官︵animal organ︶と呼ぶ。古くから人体生理学において、栄養・成長・生殖・呼吸・血液循環・排出などの植物性機能に対し、生体の対外的・能動的働きかけとしての行動系を実現することが多いため、﹁動物﹂の名を冠し呼ばれる。植物でも動物性機能は多く見られるが、医学では現在でも用いられている。
動物極︵animal pole︶
動物の卵細胞や初期胚において、極体の生じる極、または重力と平衡な環境において上方に位置する極を指す。これらは一致しないこともある。この極の付近から上記の動物性器官︵神経系・感覚器官・運動器官︶が生じると考えられたためこの名があるが、そうでない場合もある。
(一)^ Shalchian-Tabrizi et al., 2008
(二)^ abcdefg巌佐ほか 2013, p. 994e.
(三)^ ab"動物". デジタル大辞泉︵小学館︶. コトバンクより2018年7月18日閲覧。
(四)^ 八杉貞夫 (2018), “動物学の歴史 ―2000年の動物学史のエッセンス”, p. 2 in日本動物学会 2018
(五)^ 藤田 2010, p. 91.
(六)^ Lisa A. Urry; Michael L. Cain; Steven A. Wasserman; Peter V. Minorsky; Jane B. Reece 池内昌彦、伊藤元己、箸本春樹 、道上達男訳 (2018-03-20). キャンベル生物学 原書11版. 丸善出版. p. 655. ISBN 978-4621302767
(七)^ P. レーヴン; J. ロソス; S. シンガー; G. ジョンソン (2007-05-01). レーヴン ジョンソン 生物学︿下﹀︵原書第7版︶. 培風館. p. 518
(八)^ “中学校理科教科書﹁未来へ広がるサイエンス﹂”. 啓林館. 2018年7月11日閲覧。
(九)^ abcdeAdl et al. 2019, pp. 4–119.
(十)^ ab矢﨑裕規・島野智之 (2020). “真核生物の高次分類体系の改訂 ―Adl et al. (2019) について―”. タクサ 48: 71–83.
(11)^ abcdefghijklmnopqrstuvwxyzaaabacad角井敬知 (2018), “動物界の分類群・系統 ―いまだに解けない古い関係”, pp. 54–57 in日本動物学会 2018
(12)^ 藤田 2010, p. 99.
(13)^ Ruggiero, Michael A.; Gordon, Dennis P.; Orrell, Thomas M.; Bailly, Nicolas; Bourgoin, Thierry; Brusca, Richard C.; Cavalier-Smith, Thomas; Guiry, Michael D. et al.. “A Higher Level Classification of All Living Organisms”. PLoS ONE 10(4): 1–60. doi:10.1371/journal.pone.0119248.
(14)^ Tedersoo, Leho; Sánchez-Ramírez, Santiago; Kõljalg, Urmas; Bahram, Mohammad; DÖring, Markus; Schigel, Dmitry; May, Tom; Ryberg, Martin et al. (2018). “High-level classification of the Fungi and a tool for evolutionary ecological analyses”. Fungal Diversity 90: 135–159. doi:10.1007/s13225-018-0401-0.
(15)^ 巌佐ほか 2013, p. 1552.
(16)^ 動物命名法国際審議会 2005, 条1.
(17)^ 動物命名法国際審議会 2005, 表紙.
(18)^ 動物命名法国際審議会 2005, 条1.1.1.
(19)^ 動物命名法国際審議会 2005, 用語集.
(20)^ 中野隆文 (2018), “種と学名,高次分類群 ―動物の名称と名称に関するルール”, pp. 46–47 in日本動物学会 2018
(21)^ Avila, Vernon L. (1995). Biology: Investigating Life on Earth. Jones & Bartlett Learning. pp. 767–. ISBN 978-0-86720-942-6. https://books.google.com/books?id=B_OOazzGefEC&pg=PA767
(22)^ ab“Palaeos:Metazoa”. Palaeos. 2018年2月25日閲覧。
(23)^ Bergman, Jennifer. “Heterotrophs”. 2007年8月29日時点のオリジナルよりアーカイブ。2007年9月30日閲覧。
(24)^ Mentel, Marek; Martin, William (2010). “Anaerobic animals from an ancient, anoxic ecological niche”. BMC Biology 8: 32. doi:10.1186/1741-7007-8-32. PMC 2859860. PMID 20370917. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2859860/.
(25)^ Saupe, S. G.. “Concepts of Biology”. 2007年9月30日閲覧。
(26)^ Minkoff, Eli C. (2008). Barron's EZ-101 Study Keys Series: Biology (2nd, revised ed.). Barron's Educational Series. p. 48. ISBN 978-0-7641-3920-8
(27)^ 白山義久 (2000), “総合的観点から見た無脊椎動物の多様性と系統︵1-3-2 動物界の門レベルの多様性︶”, pp. 14–25 in白山 2000
(28)^ abcdefghi藤田 2010, pp. 102–106.
(29)^ 浅島・駒崎 2011, p. 49.
(30)^ abcdef濱田博司 (2018), “左右軸形成 ―なぜ心臓や胃は左に?”, pp. 308-309 in日本動物学会 2018
(31)^ abc巌佐ほか 2013, p. 1401.
(32)^ abcdefghi松尾勲 (2018), “頭尾軸・背腹軸形成 ―動物界に共通する普遍的な体制”, pp. 304-307 in日本動物学会 2018
(33)^ ab佐藤ほか 2004, pp. 30–37.
(34)^ abcd佐藤ほか 2004, pp. 38–41.
(35)^ abcdefg白山義久 (2000), “総合的観点から見た無脊椎動物の多様性と系統︵1-3-2 動物界の門レベルの多様性︶”, pp. 16–17 in白山 2000
(36)^ abcdefghijklmnopqr久米・團 1957, pp. 35–37︵総説︶
(37)^ abcd佐藤ほか 2004, p. 11.
(38)^ abc駒崎伸二・浅島誠 (2018), “胚葉形成 ―動物の体をつくる基本作業”, pp. 296–299 in日本動物学会 2018
(39)^ abcdefg白山義久 (2000), “総合的観点から見た無脊椎動物の多様性と系統︵1-3-2 動物界の門レベルの多様性︶”, pp. 19-20 in白山 2000
(40)^ 巌佐ほか 2013, p. 848.
(41)^ abKozloff 1990, pp. 7–8.
(42)^ ab巌佐ほか 2013, p. 405.
(43)^ 松本信二、船越浩海、玉野井逸朗﹃細胞の増殖と生体システム﹄︵初版︶学会出版センター、1993年。ISBN 4-7622-6737-6。 pp.47-50、3.細胞の微細構造とその機能、3.1.細胞と膜
(44)^ 武村政春﹁第1章 エピジェネティクスを理解するための基礎知識、1-1 DNAとセントラルドグマ﹂﹃DNAを操る分子たち﹄︵初版第1刷︶技術評論社、2012年、14–24頁。ISBN 978-4-7741-4998-1。
(45)^ 松本信二、船越浩海、玉野井逸朗﹁3.細胞の微細構造とその機能、3.3.真核生物、3.3.1真核生物の構造と機能概説﹂﹃細胞の増殖と生体システム﹄︵初版︶学会出版センター、1993年、53–56頁。ISBN 4-7622-6737-6。
(46)^ 井出利憲﹁第6章 テロメアとは何か﹂﹃細胞の運命Ⅳ細胞の老化﹄︵初版︶サイエンス社、2006年、65–75頁。ISBN 4-7819-1127-7。
(47)^ ﹁︻細胞骨格タンパク質︼﹂﹃生化学辞典第2版﹄︵第2版第6刷︶東京化学同人、1995年、534頁。ISBN 4-8079-0340-3。
(48)^ 林純一﹁ミトコンドリアDNAに突然変異をもつ細胞は自然免疫により排除されることを発見﹂筑波大学生命科学研究科発表 ﹁Journal of Experimental Medicine﹂電子版 2011.Oct.12
(49)^ 黒岩常祥﹃ミトコンドリアはどこからきたか﹄日本放送出版、2000年6月30日第1刷発行。ISBN 4140018879。
(50)^ Alberts, Bruce; Johnson, Alexander; Lewis, Julian; Raff, Martin; Roberts, Keith; Walter, Peter (2002). Molecular Biology of the Cell (4th ed.). Garland Science. ISBN 0-8153-3218-1. https://www.ncbi.nlm.nih.gov/books/NBK26810/
(51)^ Sangwal, Keshra (2007). Additives and crystallization processes: from fundamentals to applications. John Wiley and Sons. p. 212. ISBN 978-0-470-06153-4
(52)^ Magloire, Kim (2004). Cracking the AP Biology Exam, 2004–2005 Edition. The Princeton Review. p. 45. ISBN 978-0-375-76393-9
(53)^ Starr, Cecie (2007-09-25). Biology: Concepts and Applications without Physiology. Cengage Learning. pp. 362, 365. ISBN 0495381500. https://books.google.com/?id=EXNFwB-O-WUC&pg=PA362
(54)^ Knobil, Ernst (1998). Encyclopedia of reproduction, Volume 1. Academic Press. p. 315. ISBN 978-0-12-227020-8
(55)^ abcdefghijklmn小林一也 (2018), “有性生殖と無性生殖 ―生殖戦略の多様性”, pp. 274–275 in日本動物学会 2018
(56)^ Hamilton, Matthew B. (2009). Population genetics. Wiley-Blackwell. p. 55. ISBN 978-1-4051-3277-0
(57)^ ab巌佐ほか 2013, p. 638.
(58)^ abc巌佐ほか 2013, p. 744.
(59)^ 巌佐ほか 2013, pp. 1105–1106.
(60)^ abc浅島・駒崎 2011, p. 33.
(61)^ abcde浅島誠・駒崎伸二 (2018), “さまざまな動物の発生 ―卵から形づくりの始まり”, pp. 270–273 in日本動物学会 2018
(62)^ 巌佐ほか 2013, p. 1313.
(63)^ ab久米・團 1957, p. 371︵総説︶
(64)^ 巌佐ほか 2013, p. 1430.
(65)^ abcdefghijklmKozloff 1990, pp. 4–5.
(66)^ ab巌佐ほか 2013, p. 406.
(67)^ ab久米・團 1957, pp. 39–40︵総説︶
(68)^ 巌佐ほか 2013, p. 1397.
(69)^ 巌佐ほか 2013, p. 923.
(70)^ abcdefgh久米・團 1957, pp. 28–31︵総説︶
(71)^ abcde巌佐ほか 2013, p. 1443.
(72)^ abcdefghijklKozloff 1990, p. 3.
(73)^ abcdef久米・團 1957, p. 5︵総説︶
(74)^ abcdefg久米・團 1957, pp. 31–33︵総説︶
(75)^ abcdefghijklmnopqrMartín-Durán, José M.; Marlétaz, Ferdinand (2020). “Unravelling spiral cleavage”. Development 147: 1–7. doi:10.1242/dev.181081.
(76)^ abcd上野秀一 (2018), “卵割 ―大きな卵はなぜ速く分裂するのか”, pp. 294–295 in日本動物学会 2018
(77)^ abcdefghijklmnopqrstuvwx久米・團 1957, pp. 33–35︵総説︶
(78)^ 浅島・駒崎 2011, p. 39.
(79)^ 浅島・駒崎 2011, p. 42.
(80)^ abKozloff 1990, pp. 5–7.
(81)^ ab浅島・駒崎 2011, p. 126.
(82)^ ab浅島・駒崎 2011, p. 108.
(83)^ ab浅島・駒崎 2011, pp. 72–73.
(84)^ abc浅島・駒崎 2011, p. 115.
(85)^ abc浅島・駒崎 2011, p. 110.
(86)^ 浅島・駒崎 2011, pp. 115–117.
(87)^ 浅島・駒崎 2011, p. 119.
(88)^ abc藤田 2010, pp. 99–101.
(89)^ abc古屋秀隆 (2000), “後生動物の起源”, pp. 106–107 in白山 2000
(90)^ abcdefghijdos Reis et al. 2015, pp. 2939–2950.
(91)^ ab藤田 2010, p. 92.
(92)^ 土屋 2013, pp. 11–12.
(93)^ Erwin, D.H.; Laflamme, M.; Tweedt, S.M.; Sperling, E.A.; Pisani, D.; Peterson, K.J. (2011). “The Cambrian conundrum: early divergence and later ecological success in the early history of animals”. Science 334: 1091–1097.
(94)^ Budd, G.E. (2008). “The earliest fossil record of the animals and its significance”. Philos. Trans. R. Soc. Lond. B Biol. Sci. 363: 1425–1434.
(95)^ abMaloof, A.C.; Porter, S.M.; Moore, J.L.; Dudas, F.O.; Bowring, S.A.; Higgins, J.A.; Fike, D.A.; Eddy, M.P. (2010). “The earliest Cambrian record of animals and ocean geochemical change”. Geol. Soc. Am. Bull. 122 (11–12): 1731–1774. Bibcode: 2010GSAB..122.1731M. doi:10.1130/B30346.1.
(96)^ abcd土屋 2013, pp. 12–13.
(97)^ abcBrain, C.K.; Prave, Anthony R.; Hoffmann, Karl-Heinz; Fallick, Anthony E.; Botha, Andre; Herd, Donald A.; Sturrock, Craig; Young, Iain et al. (2012). “The first animals: ca. 760-million-year-old sponge-like fossils from Namibia”. S Afr J Sci. 108 (1/2): 1–8. doi:10.4102/sajs.v108i1/2.658.
(98)^ ﹃ナショナル ジオグラフィック にわかには信じがたい本当にあったこと﹄日経ナショナル ジオグラフィック、2019年3月25日、299頁。
(99)^ 松本 2015, p. 3.
(100)^ Maloof, Adam C.; Rose, Catherine V.; Beach, Robert; Samuels, Bradley M.; Calmet, Claire C.; Erwin, Douglas H.; Poirier, Gerald R.; Yao, Nan et al. (17 August 2010). “Possible animal-body fossils in pre-Marinoan limestones from South Australia”. Nature Geoscience 3(9): 653–659. Bibcode: 2010NatGe...3..653M. doi:10.1038/ngeo934. http://www.nature.com/ngeo/journal/v3/n9/full/ngeo934.html.
(101)^ Love, G.D.; Grosjean, E.; Stalvies, C.; Fike, D.A.; Grotzinger, J.P.; Bradley, A.S.; Kelly, A.E.; Bhatia, M. et al. (2009). “Fossil steroids record the appearance of Demospongiae during the Cryogenian period”. Nature 457: 718–721. doi:10.1038/nature0767.
(102)^ Siegl, A.; Kamke, J.; Hochmuth, T.; Piel, J.; Richter, M.; Liang, C.; Dandekar, T.; Hentschel, U. (2011). “Single-cell genomics reveals the lifestyle of Poribacteria, a candidate phylum symbiotically associated with marine sponges”. ISME J. 5: 61–70.
(103)^ 土屋 2013, pp. 13–18.
(104)^ Xiao, S.; Zhang, Y.; Knol (1998). “Three-dimensional preservation of algae and animal embryos in a Neoproterozoic phosphorite”. Nature 391: 553–558. doi:10.1038/35318.
(105)^ Butterfield, N.J.. “Paleontology. Terminal developments in Ediacaran embryology”. Science 334: 1655–1656.
(106)^ Huldtgren, T.; Cunningham, J.A.; Yin, C.; Stampanoni, M.; Marone, F.; Donoghue, P.C.J.; Bengtson, S. (2011). “Fossilized nuclei and germination structures identify Ediacaran "animal embryos" as encysting protists”. Science 334: 1696–1699.
(107)^ Zhang, X.-G.; Pratt, B.R. (2014). “Possible algal origin and life cycle of Ediacaran Doushantuo microfossils with dextral spiral structure”. J. Paleontol. 88: 92–98.
(108)^ 土屋 2013, p. 164.
(109)^ abcDunn, Frances S.; Liu, Alexander G.; Donoghue, Philip C. J. (2018). “Ediacaran developmental biology”. Biol. Rev. 93: 914–932. doi:10.1111/brv.12379.
(110)^ abc土屋 2013, pp. 21–40.
(111)^ 土屋 2013, p. 28.
(112)^ Two Explosive Evolutionary Events Shaped Early History Of Multicellular Life
(113)^ Shen, Bing; Dong, Lin; Xiao, Shuhai; Kowalewski, Michał (2008). “The Avalon Explosion: Evolution of Ediacara Morphospace”. Science 319 (5859): 81–84. Bibcode: 2008Sci...319...81S. doi:10.1126/science.1150279. PMID 18174439. http://www.sciencemag.org/content/319/5859/81.short.
(114)^ Yin, Z.; Zhu, M.; Davidson, E.H.; Bottjer, D.J.; Zhao, F.; Tafforeau, P. (2015). “Sponge grade body fossil with cellular resolution dating 60 Myrbefore the Cambrian”. Proc. Natl. Acad. Sci. USA 112: E1453–E1460. doi:10.1073/pnas.1414577112.
(115)^ Antcliffe, J.B.; Callow, R.H.; Brasier, M.D. (2014). “Giving the early fossil record of sponges a squeeze”. Biol. Rev. Camb. Philos. Soc. 89: 972–1004.
(116)^ Fedonkin, M.A.; Simonetta, A.; Ivantsov, A.Y. (2007). “New data on Kimberella, the Vendian mollusc-like organism (White Sea region, Russia): palaeoecological and evolutionary implications”. Geol. Soc. Lond. Spec. Publ. 286: 157–179.
(117)^ Liu, A.G.; Matthews, J.J.; Menon, L.R.; McIlroy, D.; Brasier, M.D. (2014). “Haootia quadriformis n. gen., n. sp., interpreted as a muscular cnidarian impression from the Late Ediacaran period (approx. 560 Ma)”. Proc. Biol. Sci. 281, 20141202. doi:10.1098/rspb.2014.1202.
(118)^ 土屋 2013, pp. 33–35.
(119)^ 土屋 2013, pp. 35–36.
(120)^ Carbone, C.; Narbonne, G.M. (2014). “When life got smart: the evolution of behavioral complexity through the Ediacaran and Early Cambrian of NW Canada”. J. Paleontol. 88: 309–330.
(121)^ Mángano, M.G.; Buatois, L.A. (2014). “Decoupling of body-plan diversification and ecological structuring during the Ediacaran-Cambrian transition: evolutionary and geobiological feedbacks”. Proc. Biol. Sci. 281, 20140038.
(122)^ Liu, A.G.; Mcllroy, D.; Brasier, M.D. (2010). “First evidence for locomotion in the Ediacara biota from the 565 Ma Mistaken Point Formation, Newfoundland”. Geology 38.
(123)^ Rogov, V.I.; Marusin, V.; Bykova, N.; Goy, Y.; Nagovitsin, K.E.; Kochnev, B.B.; Karlova, G.A.; Grazhdankin, D. (2012). “The oldest evidence of bioturbation on Earth”. Geology 40: 395–398.
(124)^ Pecoits, E.; Konhauser, K.O.; Aubet, N.R.; Heaman, L.M.; Veroslavsky, G.; Stern, R.A.; Gingras, M.K. (2012). “Bilaterian burrows and grazing behavior at >585 million years ago”. Science 336: 1693–1696.
(125)^ 土屋 2013, pp. 165–166.
(126)^ 土屋 2013, pp. 166–167.
(127)^ abcdefghijklmno藤田 2010, pp. 92–98.
(128)^ 土屋 2013, pp. 169–171.
(129)^ ab“New Timeline for Appearances of Skeletal Animals in Fossil Record Developed by UCSB Researchers”. The Regents of the University of California (2010年11月10日). 2021年8月28日閲覧。
(130)^ 土屋 2013, pp. 164–165.
(131)^ abcd土屋 2013, pp. 171–174.
(132)^ Conway-Morris 2003, pp. 505–515.
(133)^ 土屋 2013, pp. 179–181.
(134)^ Valentine, JW; Jablonski, D; Erwin, DH (1999). “Fossils, molecules and embryos: new perspectives on the Cambrian explosion”. Development 126 (5): 851–9. PMID 9927587. http://dev.biologists.org/content/126/5/851.long.
(135)^ Budd, Graham (2013). “At the origin of animals: the revolutionary cambrian fossil record”. Current Genomics 14(6): 344–354. doi:10.2174/13892029113149990011. PMC 3861885. PMID 24396267. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3861885/.
(136)^ Erwin, D. H.; Laflamme, M.; Tweedt, S. M.; Sperling, E. A.; Pisani, D.; Peterson, K. J. (2011). “The Cambrian conundrum: early divergence and later ecological success in the early history of animals”. Science 334 (6059): 1091–1097. Bibcode: 2011Sci...334.1091E. doi:10.1126/science.1206375. PMID 22116879.
(137)^ Kouchinsky, A.; Bengtson, S.; Runnegar, B. N.; Skovsted, C. B.; Steiner, M.; Vendrasco, M. J. (2012). “Chronology of early Cambrian biomineralization”. Geological Magazine 149 (2): 221–251. Bibcode: 2012GeoM..149..221K. doi:10.1017/s0016756811000720.
(138)^ Servais, T.; Harper, D.A.T. (2018). “The Great Ordovician Biodiversification Event (GOBE): definition, concept and duration”. Lethaia 51: 151–164.
(139)^ García-Bellido, Diego C; Paterson, John R (2014). “A new vetulicolian from Australia and its bearing on the chordate affinities of an enigmatic Cambrian group”. BMC Evolutionary Biology 14: 214. doi:10.1186/s12862-014-0214-z. PMC 4203957. PMID 25273382. http://www.biomedcentral.com/1471-2148/14/214/abstract#.
(140)^ abSmith, Martin R.; Ortega-Hernández, Javier (2014-08-17). “Hallucigenia’s onychophoran-like claws and the case for Tactopoda” (英語). Nature 514 (7522): 363–366. doi:10.1038/nature13576. ISSN 0028-0836. https://doi.org/10.1038/nature13576.
(141)^ abcHernández, Javier Ortega (英語). Lobopodians. http://www.academia.edu/16933971/Lobopodians.
(142)^ abcde白山義久・久保田信・駒井智幸・西川輝昭・月井雄二・加藤哲哉・窪寺恒己・齋藤寛・長谷川和範・藤田敏彦・土田真二 (2005-03-20). 水の生物. 小学館の図鑑 NEO. ISBN 4092172079
(143)^ 藤田 2010, p. 127.
(144)^ abc藤田 2010, pp. 119–120.
(145)^ abcde中野裕昭 (2018), “珍無腸形動物 ―左右相称動物の祖先に迫る?”, pp. 86–87 in日本動物学会 2018
(146)^ abcdeCannon, Johanna T.; Vellutini, Bruno C.; Smith III, Julian.; Ronquist, Frederik; Jondelius, Ulf; Hejnol, Andreas (3 February 2016). “Xenacoelomorpha is the sister group to Nephrozoa”. Nature 530 (7588): 89–93. Bibcode: 2016Natur.530...89C. doi:10.1038/nature16520. PMID 26842059. http://www.nature.com/nature/journal/v530/n7588/full/nature16520.html 2016年2月3日閲覧。.
(147)^ abcRouse, Greg W.; Wilson, Nerida G.; Carvajal, Jose I.; Vrijenhoek, Robert C. (2016-02). “New deep-sea species of Xenoturbella and the position of Xenacoelomorpha” (英語). Nature 530 (7588): 94–97. doi:10.1038/nature16545. ISSN 0028-0836. http://www.nature.com/articles/nature16545.
(148)^ abcPhilippe, H.; Brinkmann, H.; Copley, R. R.; Moroz, L. L.; Nakano, H.; Poustka, A. J.; Wallberg, A.; Peterson, K. J. et al. (2011). “Acoelomorph flatworms are deuterostomes related to Xenoturbella”. Nature 470 (7333): 255–258. doi:10.1038/nature09676. PMC 4025995. PMID 21307940. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4025995/.
(149)^ abcdePhilippe, H.; Poustka, Albert J.; Chiodin, Marta; J.Hoff, Katharina; Dessimoz, Christophe; Tomiczek, Bartlomiej; Schiffer, Philipp H.; Müller, Steven et al. (2019). “Mitigating Anticipated Effects of Systematic Errors Supports Sister-Group Relationship between Xenacoelomorpha and Ambulacraria”. Current Biology 29(11): 1818–1826. doi:10.1016/j.cub.2019.04.009.
(150)^ 藤田 2010, p. 124.
(151)^ abc藤田 2010, p. 113.
(152)^ abc藤田 2010, p. 122.
(153)^ abcdefghi柁原宏 (2018), “腹毛動物・扁形動物・顎口動物・微顎動物・輪形動物・紐形動物 ―人目に触れないマイナー分類群”, pp. 62-63 in日本動物学会 2018
(154)^ 田中正敦 (2018), “環形動物︵有鬚動物・ユムシ・星口動物を含む︶ ―誤解されていた系統関係”, pp. 70–71 in日本動物学会 2018
(155)^ 藤田 2010, p. 106.
(156)^ Dunn, Casey W.; Hejnol, Andreas; Matus, David Q.; Pang, Kevin; Browne, William E.; Smith, Stephen A.; Seaver, Elaine; Rouse, Greg W. et al. (2008-03-05). “Broad phylogenomic sampling improves resolution of the animal tree of life” (英語). Nature 452 (7188): 745–749. doi:10.1038/nature06614. ISSN 0028-0836. http://www.nature.com/doifinder/10.1038/nature06614.
(157)^ Egger, Bernhard; Steinke, Dirk; Tarui, Hiroshi; Mulder, Katrien De; Arendt, Detlev; Borgonie, Gaëtan; Funayama, Noriko; Gschwentner, Robert et al. (2009-05-11). “To Be or Not to Be a Flatworm: The Acoel Controversy” (英語). PLOS ONE 4(5): e5502. doi:10.1371/journal.pone.0005502. ISSN 1932-6203. PMC 2676513. PMID 19430533. http://journals.plos.org/plosone/article?id=10.1371/journal.pone.0005502.
(158)^ abcdefghiGiribet 2016, pp. 14–21
(159)^ abcdefghijklmLaumer et al. 2019, pp. 1–10
(160)^ abcdLu, Tsai-Ming; Kanda, Miyuki; Satoh, Noriyuki; Furuya, Hidetaka (2017). “The phylogenetic position of dicyemid mesozoans offers insights into spiralian evolution”. Zoological Letters 3(6): 1–9. doi:10.1186/s40851-017-0068-5.
(161)^ abSchiffer, Philipp H.; Robertson, Helen E.; Telford, Maximilian J. (2018). “Orthonectids Are Highly Degenerate Annelid Worms”. Current Biology 28(12): 1970–1974. doi:10.1016/j.cub.2018.04.088.
(162)^ abc藤田 2010, p. 114.
(163)^ Pisani, Davide; Pett, Walker; Dohrmann, Martin; Feuda, Roberto; Rota-Stabelli, Omar; Philippe, Hervé; Lartillot, Nicolas; Wörheide, Gert (15 December 2015). “Genomic data do not support comb jellies as the sister group to all other animals”. Proceedings of the National Academy of Sciences 112 (50): 15402–15407. doi:10.1073/pnas.1518127112. PMC 4687580. PMID 26621703. http://www.pnas.org/content/112/50/15402.
(164)^ Simion, Paul; Philippe, Hervé; Baurain, Denis; Jager, Muriel; Richter, Daniel J.; Franco, Arnaud Di; Roure, Béatrice; Satoh, Nori et al. (3 April 2017). “A Large and Consistent Phylogenomic Dataset Supports Sponges as the Sister Group to All Other Animals”. Current Biology 27(7): 958–967. doi:10.1016/j.cub.2017.02.031. PMID 28318975. https://doi.org/10.1016/j.cub.2017.02.031.
(165)^ Feuda, Roberto; Dohrmann, Martin; Pett, Walker; Philippe, Hervé; Rota-Stabelli, Omar; Lartillot, Nicolas; Wörheide, Gert; Pisani, Davide (2017). “Improved Modeling of Compositional Heterogeneity Supports Sponges as Sister to All Other Animals”. Current Biology 27(24): 3864. doi:10.1016/j.cub.2017.11.008. PMID 29199080. http://linkinghub.elsevier.com/retrieve/pii/S0960982217314537.
(166)^ abLaumer, Christopher E.; Gruber-Vodicka, Harald; Hadfield, Michael G.; Pearse, Vicki B.; Riesgo, Ana; Marioni, John C.; Giribet, Gonzalo (2018). “Support for a clade of Placozoa and Cnidaria in genes with minimal compositional bias”. eLIFE 7:e36278: 1–19. doi:10.7554/eLife.36278.
(167)^ abDunn, Casey W.; Hejnol, Andreas; Matus, David Q.; Pang, Kevin; Browne, William E.; Smith, Stephen A.; Seaver, Elaine; Rouse, Greg W. et al. (2008). “Broad phylogenomic sampling improves resolution of the animal tree of life”. Nature 452 (7188): 745–749. Bibcode: 2008Natur.452..745D. doi:10.1038/nature06614. PMID 18322464. http://www.nature.com/doifinder/10.1038/nature06614.
(168)^ abcdHenjol, Andreas; Matthias, Obst; Stamatakis, Alexandros; Ott, Michael; Rouse, Greg W.; Edgecombe, Gregory D.; Martinez, Pedro; Jaume, Baguñà et al. (2009). “Assessing the root of bilaterian animals with scalable phylogenomic methods”. Proc. R. Soc. B 276: 4261–4270. doi:10.1098/rspb.2009.0896.
(169)^ Whelan, Nathan V.; Kocot, Kevin M.; Moroz, Leonid L.; Halanych, Kenneth M. (2015). “Error, signal, and the placement of Ctenophora sister to all other animals”. PNAS 112 (18): 5773–5778. doi:10.1073/pnas.1503453112.
(170)^ Whelan, Nathan V.; Kocot, Kevin M.; Moroz, Tatiana P.; Mukherjee, Krishanu; Williams, Peter; Paulay, Gustav; Moroz, Leonid L.; Halanych, Kenneth M. (2017). “Ctenophore relationships and their placement as the sister group to all other animals”. Nature Ecology & Evolution 1(11): 1737. doi:10.1038/s41559-017-0331-3. http://www.nature.com/articles/s41559-017-0331-3.
(171)^ Wainright, Patricia O.; Hinkle, Gregory; Sogin, Mitchell L.; Stickel, Shawn K. (1993). “Monophyletic Origins of the Metazoa: An Evolutionary Link with Fungi”. Science New Series 260 (5106): 340–342.
(172)^ Jessop, Nancy Meyer (1970). Biosphere; a study of life. Prentice-Hall. p. 428
(173)^ Sumich, James L. (2008). Laboratory and Field Investigations in Marine Life. Jones & Bartlett Learning. p. 67. ISBN 978-0-7637-5730-4
(174)^ abc藤田 2010, pp. 117–120.
(175)^ Safra, Jacob E. (2003). The New Encyclopædia Britannica, Volume 16. Encyclopædia Britannica. p. 523. ISBN 978-0-85229-961-6
(176)^ abcde中野裕昭 (2018), “刺胞動物・有櫛動物・平板動物・海綿動物 ―左右相称でない動物たち”, pp. 58–59 in日本動物学会 2018
(177)^ 藤田 2010, pp. 120–121.
(178)^ abc久保田信 (2000), 有櫛動物と刺胞動物の関係, pp. 116-117 in白山 2000
(179)^ abc藤田 2010, pp. 122–132.
(180)^ abMinelli, Alessandro (2009). Perspectives in Animal Phylogeny and Evolution. Oxford University Press. p. 53. ISBN 978-0-19-856620-5. https://books.google.com/books?id=jIASDAAAQBAJ&pg=PA53
(181)^ abcBrusca, Richard C. (2016). Introduction to the Bilateria and the Phylum Xenacoelomorpha | Triploblasty and Bilateral Symmetry Provide New Avenues for Animal Radiation. Sinauer Associates. pp. 345–372. ISBN 978-1605353753. http://www.sinauer.com/media/wysiwyg/samples/Brusca3e_Chapter_9.pdf
(182)^ Quillin, K. J. (May 1998). “Ontogenetic scaling of hydrostatic skeletons: geometric, static stress and dynamic stress scaling of the earthworm lumbricus terrestris”. The Journal of Experimental Biology 201 (12): 1871–83. PMID 9600869. https://journals.biologists.com/jeb/cgi/pmidlookup.
(183)^ Westblad, E. (1949). “Xenoturbella bocki n. g., n. sp., a peculiar, primitive Turbellarian type”. Arkiv för Zoologi 1: 3–29.
(184)^ Bourlat, S. et al. (2006). “Deuterostome phylogeny reveals monophyletic chordates and the new phylum Xenoturbellida”. Nature 444: 85–88.
(185)^ Perseke, M.; Hankeln, T.; Weich, B.; Fritzsch, G.; Stadler, P.F.; Israelsson, O.; Bernhard, D.; Schlegel, M. (August 2007). “The mitochondrial DNA of Xenoturbella bocki: genomic architecture and phylogenetic analysis”. Theory Biosci 126 (1): 35–42. doi:10.1007/s12064-007-0007-7. PMID 18087755. http://www.bioinf.uni-leipzig.de/Publications/PREPRINTS/07-009.pdf.
(186)^ Baguñà, J; Riutort, M (2004). “Molecular phylogeny of the Platyhelminthes”. Can J Zool 82: 168–193.
(187)^ Nakano, H.; Lundin, K.; Bourlat, S.J.; Telford, M.J. (2013). “Xenoturbella bocki exhibits direct development with similarities to Acoelomorpha”. Nature Communications 4(1): 1537. doi:10.1038/ncomms2556.
(188)^ 馬渡 2013, pp. 27–29.
(189)^ 筑波大学﹃珍渦虫はもともと単純か複雑か ―まだ続く珍無腸動物門の系統樹上の放浪―﹄︵プレスリリース︶2019年5月27日。https://www.tsukuba.ac.jp/journal/images/pdf/190527nakano-2.pdf。2021年8月20日閲覧。
(190)^ 後藤太一郎 (2000), “31. 毛顎動物門 Phylum CHAETOGNATHA”, pp. 235–237 in白山 2000
(191)^ abcdefg後藤太一郎 (2018), “毛顎動物 ―謎に包まれた系統的位置”, pp. 84–85 in日本動物学会 2018
(192)^ Telford, Maximilian J.; Holland, P. W. H. (1993). “The Phylogenetic Affinities of the Chaetognaths: A Molecular Analysis”. Mol. Biol. Evol. 10(3): 660–676.
(193)^ Wada, Hiroshi; Satoh, Noriyuki. “Details of the evolutionary history from invertebrates to vertebrates, as deduced from the sequences of 18S rDNA”. Proc. Natl. Acad. Sci. 91(5): 1801–1804. doi:10.1073/pnas.91.5.1801.
(194)^ abcdMallatt, Jon (2010). “Nearly complete rRNA genes assembled from across the metazoan animals: Effects of more taxa, a structure-based alignment, and paired-sites evolutionary models on phylogeny reconstruction”. Molecular Phylogenetics and Evolution 55: 1–17. doi:10.1016/j.ympev.2009.09.028.
(195)^ ab藤田 2010, pp. 122–123.
(196)^ ab藤田 2010, p. 108.
(197)^ Dawkins, Richard (2005). The Ancestor's Tale: A Pilgrimage to the Dawn of Evolution. Houghton Mifflin Harcourt. p. 381. ISBN 978-0-618-61916-0
(198)^ abc藤田 2010, p. 150.
(199)^ abcde嶋田大輔 (2018), “線形動物・類線形動物 ―昆虫に匹敵する多様性の持ち主?”, pp. 72–73 in日本動物学会 2018
(200)^ 藤田 2010, pp. 150–152.
(201)^ ab藤田 2010, p. 153.
(202)^ ab山崎博史 (2018), “鰓曳動物・胴甲動物・動吻動吻 ―棘に覆われた頭部をもつ動物たち”, pp. 74–75 in日本動物学会 2018
(203)^ Miller, Stephen A.; Harley, John P. (2006). Zoology. McGraw-Hill Higher Education. p. 173. https://books.google.com/books?id=BWZFAQAAIAAJ
(204)^ Telford, Maximilian J; Bourlat, Sarah J; Economou, Andrew; Papillon, Daniel; Rota-Stabelli, Omar (2008-04-27). “The evolution of the Ecdysozoa”. Philosophical Transactions of the Royal Society B: Biological Sciences 363 (1496): 1529–1537. doi:10.1098/rstb.2007.2243. PMC 2614232. PMID 18192181. https://royalsocietypublishing.org/doi/10.1098/rstb.2007.2243.
(205)^ ab藤田 2010, pp. 155–156.
(206)^ Giribet, Gonzalo; Edgecombe, Gregory D. (2017-09-01). “Current Understanding of Ecdysozoa and its Internal Phylogenetic Relationships”. Integrative and Comparative Biology 57(3): 455–466. doi:10.1093/icb/icx072. ISSN 1540-7063. https://doi.org/10.1093/icb/icx072.
(207)^ abcdefghijGiribet, Gonzalo; Edgecombe, Gregory D. (2019-06-17). “The Phylogeny and Evolutionary History of Arthropods”. Current Biology 29(12): R592–R602. doi:10.1016/j.cub.2019.04.057. ISSN 0960-9822. http://www.sciencedirect.com/science/article/pii/S0960982219304865.
(208)^ 島野智之 (2018), “節足動物︵多足類・鋏角類︶ ―いまだ系統が解明されていない2つの大きな分類群”, pp. 78–79 in日本動物学会 2018
(209)^ abc大塚攻・田中隼人 (2020). “顎脚類︵甲殻類︶の分類と系統に関する研究の最近の動向”. タクサ 48: 49–62.
(210)^ ab藤田 2010, p. 168.
(211)^ Olesen, Jørgen; Pisani, Davide; Iliffe, Thomas M.; Legg, David A.; Palero, Ferran; Glenner, Henrik; Thomsen, Philip Francis; Vinther, Jakob et al. (2019-08-01). “Pancrustacean Evolution Illuminated by Taxon-Rich Genomic-Scale Data Sets with an Expanded Remipede Sampling” (英語). Genome Biology and Evolution 11(8): 2055–2070. doi:10.1093/gbe/evz097. https://academic.oup.com/gbe/article/11/8/2055/5528088.
(212)^ abc藤田 2010, pp. 157–158.
(213)^ Ramsköld, L.; Xianguang, Hou (1991-05). “New early Cambrian animal and onychophoran affinities of enigmatic metazoans” (英語). Nature 351 (6323): 225–228. doi:10.1038/351225a0. ISSN 0028-0836. https://www.nature.com/articles/351225a0.
(214)^ BUDD, GRAHAM E. (1996-03). “The morphology of Opabinia regalis and the reconstruction of the arthropod stem-group” (英語). Lethaia 29(1): 1–14. doi:10.1111/j.1502-3931.1996.tb01831.x. ISSN 0024-1164. https://onlinelibrary.wiley.com/doi/abs/10.1111/j.1502-3931.1996.tb01831.x.
(215)^ Budd, Graham E. (1998/ed). “The morphology and phylogenetic significance of Kerygmachela kierkegaardi Budd (Buen Formation, Lower Cambrian, N Greenland)” (英語). Earth and Environmental Science Transactions of The Royal Society of Edinburgh 89(4): 249–290. doi:10.1017/S0263593300002418. ISSN 1473-7116. https://www.cambridge.org/core/journals/earth-and-environmental-science-transactions-of-royal-society-of-edinburgh/article/abs/morphology-and-phylogenetic-significance-of-kerygmachela-kierkegaardi-budd-buen-formation-lower-cambrian-n-greenland/AF165229724342F0BD90933A037CB05F.
(216)^ Budd, Graham E. (2001-01). “Tardigrades as ‘Stem-Group Arthropods’: The Evidence from the Cambrian Fauna”. Zoologischer Anzeiger - A Journal of Comparative Zoology 240 (3–4): 265–279. doi:10.1078/0044-5231-00034. ISSN 0044-5231. https://doi.org/10.1078/0044-5231-00034.
(217)^ Jianni Liu, Degan Shu, Jian Han, Zhifei Zhang, and Xingliang Zhang (2006-06). “A large xenusiid lobopod with complex appendages from the Lower Cambrian Chengjiang Lagerstätte”. Acta Palaeontologica Polonica 51(2).
(218)^ Liu, J; Shu, D; Han, Jian; Zhang, Zhifei; Zhang, Xingliang (2007-09-26). “Morpho-anatomy of the lobopod Magadictyon cf. haikouensis from the Early Cambrian Chengjiang Lagerstätte, South China” (英語). Acta Zoologica 89(2): 183–183. doi:10.1111/j.1463-6395.2007.00307.x. https://www.researchgate.net/publication/293346517.
(219)^ Edgecombe, Gregory D. (2009-06). “Palaeontological and Molecular Evidence Linking Arthropods, Onychophorans, and other Ecdysozoa” (英語). Evolution: Education and Outreach 2(2): 178–190. doi:10.1007/s12052-009-0118-3. ISSN 1936-6434. https://evolution-outreach.biomedcentral.com/articles/10.1007/s12052-009-0118-3.
(220)^ Siveter, Derek J.; Briggs, Derek E. G.; Siveter, David J.; Sutton, Mark D.; Legg, David (2018-08-01). “A three-dimensionally preserved lobopodian from the Herefordshire (Silurian) Lagerstätte, UK” (英語). Open Science 5(8): 172101. doi:10.1098/rsos.172101. ISSN 2054-5703. http://rsos.royalsocietypublishing.org/content/5/8/172101.
(221)^ abc白山義久 (2000), “総合的観点から見た無脊椎動物の多様性と系統︵1-5-4 分子系統学︶”, pp. 34–45 in白山 2000
(222)^ abStruck, Torsten H.; Wey-Fabrizius, Alexandra R.; Golombek, Anja; Hering, Lars; Weigert, Anne; Bleidorn, Christoph; Klebow, Sabrina; Iakovenko, Nataliia et al. (2014). “Platyzoan Paraphyly Based on Phylogenomic Data Supports a Noncoelomate Ancestry of Spiralia”. Molecular Biology and Evolution 31(7): 1833-1849. doi:10.1093/molbev/msu143. PMID 24748651.
(223)^ Shankland, M.; Seaver, E. C. (2000). “Evolution of the bilaterian body plan: What have we learned from annelids?”. Proceedings of the National Academy of Sciences 97(9): 4434–7. Bibcode: 2000PNAS...97.4434S. doi:10.1073/pnas.97.9.4434. JSTOR 122407. PMC 34316. PMID 10781038. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC34316/.
(224)^ abcd藤田 2010, pp. 130–131.
(225)^ ab藤田 2010, pp. 127–128.
(226)^ Balsamo, Maria; Artois, Tom; Smith III, Julian P. S.; Todaro, M. Antonio; Guidi, Loretta (2020). “The curious and neglected soft-bodied meiofauna: Rouphozoa (Gastrotricha and Platyhelminthes)”. Hydrobiologia 847: 2613–2644.
(227)^ ab藤田 2010, pp. 135–136.
(228)^ ab藤田 2010, p. 132.
(229)^ abcde藤田 2010, pp. 136–137.
(230)^ abc佐々木猛智 (2018), “軟体動物 ―900 kgのイカ,0.01 g の巻貝”, pp. 68–69 in日本動物学会 2018
(231)^ abcdWeigert, Anne; Bleidorn, Christoph (2016). “Current status of annelid phylogeny”. Org Divers Evol 16: 345–362. doi:10.1007/s13127-016-0265-7.
(232)^ 巌佐ほか 2013, pp. 1584–1586.
(233)^ ab藤田 2010, pp. 104–105.
(234)^ ab佐藤ほか 2004, pp. 63–64.
(235)^ 白山 2000, p.23
(236)^ 白山義久 (2000), “総合的観点から見た無脊椎動物の多様性と系統︵1-3-2 動物界の門レベルの多様性︶”, pp. 21–23 in白山 2000
(237)^ ab藤田 2010, p. 169.
(238)^ ab白山義久 (2000), “総合的観点から見た無脊椎動物の多様性と系統︵1-3-1 漸進的進化思想と分子系統樹︶”, pp. 3–14 in白山 2000
(239)^ Edgecombe, Gregory D.; Giribet, Gonzalo; Dunn, Casey W.; Hejnol, Andreas; Kristensen, Reinhardt M.; Neves, Ricardo C.; Rouse, Greg W.; Worsaae, Katrine et al. (June 2011). “Higher-level metazoan relationships: recent progress and remaining questions”. Organisms, Diversity & Evolution 11(2): 151–172. doi:10.1007/s13127-011-0044-4.
(240)^ Fröbius, Andreas C.; Funch, Peter (2017-04-04). “Rotiferan Hox genes give new insights into the evolution of metazoan bodyplans”. Nature Communications 8(1). Bibcode: 2017NatCo...8....9F. doi:10.1038/s41467-017-00020-w. http://www.nature.com/articles/s41467-017-00020-w.
(241)^ Smith, Martin R.; Ortega-Hernández, Javier (2014). “Hallucigenia’s onychophoran-like claws and the case for Tactopoda”. Nature 514 (7522): 363–366. Bibcode: 2014Natur.514..363S. doi:10.1038/nature13576. https://doi.org/10.1038/nature13576.
(242)^ ab“Palaeos Metazoa: Ecdysozoa”. palaeos.com. 2017年9月2日閲覧。
(243)^ Yamasaki, Hiroshi; Fujimoto, Shinta; Miyazaki, Katsumi (June 2015). “Phylogenetic position of Loricifera inferred from nearly complete 18S and 28S rRNA gene sequences”. Zoological Letters 1: 18. doi:10.1186/s40851-015-0017-0. https://doi.org/10.1186/s40851-015-0017-0.
(244)^ Nielsen, C. (2002). Animal Evolution: Interrelationships of the Living Phyla (2nd ed.). Oxford University Press. ISBN 0-19-850682-1
(245)^ “Bilateria”. Tree of Life Web Project (2001年). 2014年8月11日閲覧。
(246)^ abcd藤田 2010, pp. 169–173.
(247)^ abc藤田敏彦 (2018), “棘皮動物 ―星形の体をもつ海のスター”, pp. 88–89 in日本動物学会 2018
(248)^ 藤田 2010, p. 173.
(249)^ 西川輝昭 (2000), “33. 半索動物 Phylum HEMICHORDATA”, pp. 253–255 in白山 2000
(250)^ abcdefghi西川輝昭 (2018), “頭索動物・尾索動物・半索動物 ―脊椎動物のルーツを探る”, pp. 90–91 in日本動物学会 2018
(251)^ ab藤田 2010, p. –173.
(252)^ ab藤田 2010, p. -173.
(253)^ ab佐藤ほか 2004, p. 117.
(254)^ abcdefg藤田 2010, pp. 174–180.
(255)^ ︵プレスリリース︶沖縄科学技術大学院大学・東邦大学、2014年9月17日。https://www.oist.jp/ja/news-center/press-releases/16643。2021年8月8日閲覧。
(256)^ Satoh, Noriyuki; Rokhsar, Daniel; Nishikawa, Teruaki (2014). “Chordate evolution and the three-phylum system”. Proceedings of Royal Society B 281 (1794): 1-10. doi:10.1098/rspb.2014.1729.
(257)^ ab甲斐嘉晃 (2018), “脊椎動物︵魚類︶ ―水中で多様に進化した分類群”, pp. 92-95 in日本動物学会 2018
(258)^ 栗田和紀 (2018), “脊椎動物︵爬虫類︶ ―陸に卵を産み始めた脊椎動物”, pp. 98-99 in日本動物学会 2018
(259)^ ab佐藤ほか 2004, p. 196.
(260)^ abc鈴木大地﹁アリストテレス﹃動物発生論﹄の現代生物学・科学哲学的検討Ⅰ : 第1巻第1章~第16章﹂﹃古典古代学﹄第6号、筑波大学大学院人文社会科学研究科古典古代学研究室、2013年、1-23頁、ISSN 1883-7352、NAID 120005373271、2021年10月1日閲覧。
(261)^ abcd松浦 2009, pp. 17–18.
(262)^ 松浦 2009, pp. 20–21.
(263)^ ab上島励 (2000), ミクソゾアの系統学的位置, p. 93 in白山 2000
(264)^ ab古屋秀隆 (2004). “中生動物研究の現状”. タクサ (日本動物分類学会) (16): 1–9.
(265)^ Kozloff 1990, pp. 212–216.
(266)^ 藤田 2010, p. 125.
(267)^ 久米又三・織田秀実 (1957), 外肛動物, pp. 171–198 in久米・團 (1957)
(268)^ 馬渡静夫﹁触手動物の系統﹂﹃哺乳類科学﹄第10巻第2号、日本哺乳類学会、1970年、61-68頁、doi:10.11238/mammalianscience.10.2_61、ISSN 0385-437X、NAID 130000884220、2021年10月1日閲覧。
(269)^ abcdefghi白山義久 (2000), “総合的観点から見た無脊椎動物の多様性と系統”, pp. 27-30 in白山 2000
(270)^ 巌佐ほか 2013, p. 1585.
(271)^ abcd藤田 2010, p. 145.
(272)^ 藤田 2010, p. 163.
(273)^ abR.S.K.バーンズ 他 著、本川達雄 監訳 訳﹃図説無脊椎動物学﹄2009年6月25日。ISBN 978-4-254-17132-7。
(274)^ 藤田 2010, p. 130.
(275)^ abcdefg正田陽一・西田恂子. "家畜". 日本大百科全書(ニッポニカ). コトバンクより2024年5月19日閲覧。
(276)^ abc巌佐ほか 2013, p. 226d.
(277)^ 正田陽一. "家禽". 改訂新版 世界大百科事典. コトバンクより2024年5月19日閲覧。
(278)^ 巌佐ほか 2013, p. 199c.
(279)^ abc“資料4﹁動物の愛護管理の歴史的変遷﹂”. 環境省. 2019年12月26日閲覧。
(280)^ "役畜". 精選版 日本国語大辞典. コトバンクより2024年5月19日閲覧。
(281)^ "使役犬". デジタル大辞泉. コトバンクより2024年5月19日閲覧。
(282)^ ab山下正男. "動物". 改訂新版 世界大百科事典. コトバンクより2024年5月19日閲覧。
(283)^ abc“愛玩動物の衛生管理の徹底に関するガイドライン2006”. 厚生労働省. 2019年12月26日閲覧。
(284)^ 川島誠一郎. "実験動物". 日本大百科全書(ニッポニカ). コトバンクより2024年5月19日閲覧。
(285)^ "疾患モデル動物". デジタル大辞泉. コトバンクより2024年5月19日閲覧。
(286)^ ab巌佐ほか 2013, p. 1398d.
(287)^ ab巌佐ほか 2013, p. 424f.
(288)^ 水野 1977, p. 266.
(289)^ abc巌佐ほか 2013, p. 995h.
(290)^ abc巌佐ほか 2013, p. 995a.
●Adl, Sina M.; Bass, David; Lane, Christopher E.; Lukeš, Julius; Schoch, Conrad L.; Smirnov, Alexey; Agatha, Sabine; Berney, Cedric et al. (2019). “Revisions to the Classification, Nomenclature, and Diversity of Eukaryote”. Journal of Eukaryotic Microbiology 66: 4-119. doi:10.1111/jeu.12691.
●Conway-Morris, S. (2003). “The Cambrian "explosion" of metazoans and molecular biology: would Darwin be satisfied?”. The International journal of developmental biology 47(7–8): 505–515. PMID 14756326. http://www.ijdb.ehu.es/web/paper.php?doi=14756326.
●Giribet, Gonzalo (2016). “Genomicas and the animal tree of life: conflicts and future prospects”. Zoologica Scripta 45(s1): 14-21. doi:10.1111/zsc.12215.
●Kozloff, Eugene N. (1990). Invertebrates. Saunders College Publishing. ISBN 0030462045
●Laumer, Christopher E.; Sørensen,Martin V. and Giribet, Gonzalo (2019). “Revisiting metazoan phylogeny with genomic sampling of all phyla”. Proc. R. Soc. B 286: 1-10. doi:10.1098/rspb.2019.0831.
●dos Reis, Mario; Thawornwattana, Yuttapong; Angelis, Konstantinos; Telford, Maximilian J.; Donoghue, Philip C.J.; Yang, Ziheng (2015). “Uncertainty in the Timing of Origin of Animals and the Limits of Precision in Molecular Timescales”. Current Biology 25: 2939-2950. doi:10.1016/j.cub.2015.09.066.
●浅島誠、駒崎伸二 著、太田次郎、赤坂甲治、浅島誠、長田敏行 編﹃動物の発生と分化﹄裳華房︿新・生命科学シリーズ﹀、2011年9月21日。ISBN 978-4785358495。
●巌佐庸、倉谷滋、斎藤成也、塚谷裕一﹃岩波生物学辞典 第5版﹄岩波書店、2013年2月26日。ISBN 9784000803144。
●岩槻邦男・馬渡峻輔監修 著、白山義久 編﹃無脊椎動物の多様性と系統﹄裳華房︿バイオディバーシティ・シリーズ﹀、2000年11月30日。ISBN 4785358289。
●久米又三、團勝磨﹃無脊椎動物発生学﹄培風館、1957年9月30日。
●佐藤矩行、野地澄晴、倉谷滋、長谷部光泰﹃発生と進化﹄岩波書店︿シリーズ 進化学﹀、2004年6月8日。ISBN 4000069241。
●土屋健 著、群馬県立自然史博物館 編﹃エディアカラ紀・カンブリア紀の生物﹄技術評論社︿生物ミステリーPRO﹀、2013年12月15日。ISBN 978-4774160849。
●動物命名法国際審議会 著、野田泰一・西川輝昭 編﹃国際動物命名規約 第4版 日本語版 ﹇追補﹈﹄日本分類学会連合、東京、2005年10月。ISBN 4-9980895-1-X。http://ujssb.org/iczn/index.html。
●公益社団法人 日本動物学会﹃動物学の百科事典﹄丸善出版、2018年9月28日。ISBN 978-4621303092。
●藤田敏彦 著、太田次郎、赤坂甲治、浅島誠、長田敏行 編﹃動物の系統分類と進化﹄裳華房︿新・生命科学シリーズ﹀、2010年4月28日。ISBN 978-4785358426。
●松浦啓一﹃動物分類学﹄東京大学出版会、2009年4月6日。ISBN 978-4130622165。
●松本忠夫 著、太田次郎、赤坂甲治、浅島誠、長田敏行 編﹃動物の生態: 脊椎動物の進化生態を中心に﹄裳華房︿新・生命科学シリーズ﹀、2015年2月21日。ISBN 978-4785358624。
●馬渡峻輔﹃動物の多様性30講﹄3巻︵初版︶、朝倉書店︿図説生物学30講︹環境編︺﹀、2013年5月25日。ISBN 978-4-254-17723-7。
●水野壽彦﹃日本淡水プランクトン図鑑﹄︵改訂︶保育社、1977年11月1日。ISBN 4586300388。
ウィキメディア・コモンズには、
動物に関連するカテゴリがあります。
ウィキスピーシーズに
動物界に関する情報があります。































































