
インフルエンザウイルス
| インフルエンザウイルス | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|

| ||||||||||||||||||
| 分類 | ||||||||||||||||||
| ||||||||||||||||||
| 種 | ||||||||||||||||||
インフルエンザウイルス︵英: influenza virus, flu virus︶は、ヒト︵人間︶に感染して、感染症であるインフルエンザを引き起こすウイルス。
ウイルスの分類上は﹁エンベロープを持つ、マイナス鎖の一本鎖RNAウイルス﹂として分類されるオルトミクソウイルス科に属する、A型インフルエンザウイルス、B型インフルエンザウイルス、C型インフルエンザウイルス、D型インフルエンザウイルスの4属4種を指す。ただし一般に﹁インフルエンザウイルス﹂と呼ぶ場合は、特にA型・B型のものを指し、その中でもさらにヒトに感染するものを意味する場合が多い︵インフルエンザワクチンはC型・D型を対象としていない︶。またヒト以外のインフルエンザウイルスは、それぞれ分離された動物の名前またはその略をつけて呼ばれるが、ヒトの場合は本項のように省略される。
本来はカモなどの水鳥を自然宿主として、その腸内に感染する弱毒性のウイルスであったものが、突然変異によってヒトの呼吸器への感染性を獲得したと考えられている。
前史[編集]
「インフルエンザの流行」も参照
近代まで[編集]
インフルエンザと人類の関わりは古く、古代エジプト時代には既にこの感染症が知られていたことが記録に残っている。ただし、インフルエンザを科学的に判定できるようになったのは20世紀以降のことで、それ以前の実績については流行特性や症例などからの推定であり、インフルエンザウイルスによるものかどうかは確かではない。そのため﹁インフルエンザ流行が記録された事例﹂も文献により様々である。
古代ギリシャのヒポクラテスの記録﹃流行病﹄︵紀元前5世紀︶にもインフルエンザではないかとみられる記述がある[1][2]。古代ローマのティトゥス・リウィウスも、震えと高熱、流行の早さといったインフルエンザの特徴を持つ病気を記録している[3]。
日本の増鏡には、元徳元年︵1329年頃︶に﹁シハブキヤミ︵咳病︶﹂が流行したとの記録が残されており、これもインフルエンザである可能性が高い[3]。
ヨーロッパでは1173年から1174年にかけてインフルエンザと思われる症状の流行が記録されており、これがインフルエンザの初めての記録とされることもある[4]。
また、1510年にはイギリスのジョンストンが、症状と、マルタ - シチリア - イタリア - スペイン - フランス - イギリスという流行経路を記録し、これがヨーロッパにおける正確な記録の最初とされる場合がある[5]。以後は記録が多くなり、特に大きかった流行としては1580年、1729-33年、1781-82年、1830-33年などが挙げられる[3]。

1918年から1919年にかけてのスペインかぜの死者数の推移 1 918年10月から11月にかけて、ニューヨーク、ロンドン、パリ、ベルリンにおいて死者が急増している
1876年のコッホによる炭疽菌の発見以降、様々な感染症についてその病原体が分離・発見されていったが、インフルエンザ病原体の発見は困難を極めた。
1889年から1900年にかけて、世界中でインフルエンザが大流行した︵ロシアかぜ︶。特に1889-1891年はヨーロッパ、アメリカで大流行した[1]。ただし、症例からみて、いくつかの型が同時に流行した可能性がある[5]。当時はまだヘマグルチニンとウイルス・ノイラミニダーゼによる区分は行われていなかったが、A型インフルエンザウイルスの内のH2N2亜型[6]、H3N8亜型[7]、H2N8亜型[3]などであったと考えられている。ただし、この流行はインフルエンザではなく、ヒトコロナウイルスOC43によるものだったという仮説も主張されている[8][9]。
1892年、北里柴三郎やリヒャルト・プファイファーがインフルエンザ患者の気道から病原体の候補となる細菌を分離し、Haemophillus influenzae︵インフルエンザ菌︶と名付けたが、コッホの原則に基づいた証明には至らなかった。実際、歴史的経緯のため今も名前が残っているが、インフルエンザ菌は、疾病﹁インフルエンザ﹂の原因ではなかった。しかし、しばしばインフルエンザに続発した呼吸器感染症の原因となる。なお、当時はまだウイルス自体が認知されておらず、ディミトリ・イワノフスキーによってウイルス︵タバコモザイクウイルス︶の存在が初めて報告されたのが、北里の発見と同じ1892年のことである。
1902年、イタリアのツェンテニ︵Centanni︶とサボヌツィ︵Savonuzzi︶は、家禽ペストの原因がウイルスであることを証明した。これは世界で4番目に発見されたウイルスだったが、ヒトと症状が異なっていたこともあり、当時はA型インフルエンザウイルスであることまでは分からなかった[4]。

スペインかぜウイルスのTEM写真
1918年から1919年にかけて発生した、スペインかぜ︵病原体はH1N1亜型)は世界的な大流行︵パンデミック︶となり、このときの感染者数は6億人、死者は4000-5000万人にのぼる︵当時の人口は20-30億人︶と言われ、第一次世界大戦終結の遠因ともいわれる。この際、候補となる細菌やウイルスが多数報告されたが、その中でも日本の山内保、坂上弘蔵、岩島寸三による1919年の報告は、インフルエンザが粘膜感染するウイルスであることを人体実験により示し[10]、後述のアンドリュースらに先立つ、最初の発見の一つとされている[11]。
1933年、イギリスのウィルソン・スミス、クリストファー・アンドリュース、パトリック・レイドローは[4]、ワシントンで発生したインフルエンザの患者から分離されたウイルスを使って、フェレットの気道に感染させてヒトのインフルエンザとよく似た症状を再現できることを実験的に示した。この実験によって、インフルエンザの病原体がウイルスであることが明らかとなり、インフルエンザウイルス︵後にA型インフルエンザウイルス︶と名付けられた。後に、この当時の流行株に対する抗体が、スペインかぜのときに採取されていた患者血清から検出され、スペインかぜの病原体がこれと同じもの︵H1N1亜型のA型インフルエンザウイルス︶であることが明らかになった[2]。
1940年、インフルエンザ患者から従来とは抗原性が異なるウイルスが分離され、B型インフルエンザウイルスと名付けられた[2]。
1946年、鼻かぜ症状を呈した患者からA、B型と異なるウイルスが分離され、1950年に病原性が証明されてC型インフルエンザウイルスと名付けられた[2]。
1955年、ドイツのヴェルナー・シェーファーは家禽ペストの原因ウイルスがA型インフルエンザウイルスであることを証明した[4]。
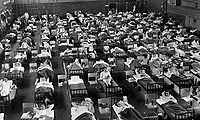
1957年のアジアかぜ流行時、病院として使われたスウェーデンの体 育館
1957年、アジアかぜとして世界的大流行を起こす。それまで流行していたH1N1亜型とは異なり、H2N2亜型に属する新型ウイルスであることが明らかになった。同時にH1N1亜型のものは姿を消した[3]。
1961年、南アフリカでアジサシが大量死しているのが見つかり、その際に野鳥から初めてインフルエンザウイルスが分離された[12]。
1965年頃、アメリカのエドウィン・キルボーンは実験レベルでウマとヒトのインフルエンザウイルスの交雑ウイルスを作ることに成功し、1971年にはブタの体内でブタとトリのインフルエンザ交雑ウイルスを作ることに成功している[12]。
1968年、香港かぜの世界的大流行。H3N2亜型に属する新型ウイルスであった。同時にH2N2亜型のものは姿を消した。現在の季節性インフルエンザの原因の1つである[3]。
1976年、アメリカ大統領のジェラルド・R・フォードはインフルエンザのパンデミックを抑えるための目的としては初めて、ワクチンの集団接種を行った。ワクチンは1億人分が用意され、4000万人に接種されたが、予想に反してインフルエンザ大流行は発生せず、接種との因果関係は不明であるものの32人にギラン・バレー症候群が発症するなどして、計画は中止された[13]。
1977年、ソ連かぜが流行。これはスペインかぜと同じH1N1亜型に属するものであった。アジアかぜ以降姿を消していたH1N1型が再び出現した理由は明らかになっていない︵一説には、アザラシなどヒト以外の生物が保存していたためとも言われている︶。このときはH3N2亜型は姿を消すことなく、以後H1N1とH3N2が毎年流行を起こすようになっている。現在の季節性インフルエンザの主な原因である[3]。
1970年代から80年代にかけて、ロバート・ウェブスターなどの研究により、インフルエンザが種の壁を乗り越えて感染する事例や、その感染経路が次々と明らかになった[13]。
1981年、ハーバード大学のドン・ウィリーらはイギリスの科学雑誌ネイチャーに、ヘマグルチニンタンパクの立体構造モデルを発表した[12][14]。
1997年、香港でH5N1亜型という高病原性鳥インフルエンザウイルスが、トリからヒトに直接感染して死者が発生した。トリからヒトへの直接感染は起きないというそれまでの定説を覆すものであり、世界的大流行が危惧されたが、ヒトの間での伝染力が低かったため大流行には至らなかった[2]。
2001年、欧米や北アフリカ、中近東の数カ国でH1N2亜型に属するウイルスがヒトの間で流行していることが確認された。これはH1N1亜型のH1とH3N2亜型のN2を併せ持ったウイルスであった。2006年現在、流行は小規模にとどまり、H1N1やH3N2に取って代わるほどの勢いはない。
2005年、ドイツ生まれのアメリカのウイルス学者ジェフェリー・タウベンバーガーは、アラスカの永久凍土に埋葬された遺体の抽出物からスペインかぜのゲノム配列を決定した[13]。さらに、河岡義裕はウィスコンシン大学において、逆遺伝学の手法を用い、スペインかぜウイルスの合成に成功し[13]、2007年にはカニクイザルに感染させることで極めて強い病原性があることも確認された[13]。
2009年メキシコから新型インフルエンザの流行が始まった。H1N1型ではあるが、スペイン風邪やソ連風邪と異なる種類である。米国では季節性インフルエンザと同程度と判断されたが、日本では﹁新型インフルエンザ﹂としての対応が取られた[15]。2011年4月からは日本でも﹁新型﹂ではなく季節性インフルエンザ﹁インフルエンザ︵H1N1)2009﹂として扱われることになった[16]。日本初の患者とされる大阪の高校生はカナダで感染して日本に持ち帰っており[13]、日本初の国内感染は神戸で確認されているが、これは診察した医師がたまたま新型インフルエンザの確認に熱心だった可能性も指摘されており、日本国内流行の発端は明らかではない[13]。
近代以降[編集]



「2009年新型インフルエンザの世界的流行」も参照
2013年3月頃から、中国上海市周辺でH7N9型インフルエンザが初めて人間に感染するようになった。4月14日現在で上海市、北京市、河南省、安徽省、江蘇省、浙江省の広い地域で51例の感染発見例がある。人間から人間への感染例は発見されていない。 2020年前半、世界的に2019新型コロナウイルスの感染が拡大。各地で都市封鎖や外出禁止令の発出が相次いだため、北半球では通常10月から5月中旬まで続くインフルエンザの流行も抑制され、欧州の流行は3月中に終息した[17]。
分類[編集]
ウイルスの分類上のインフルエンザウイルスはオルトミクソウイルス科に分類されるウイルスのうち、A型インフルエンザウイルス、B型インフルエンザウイルス、C型インフルエンザウイルスの4属4種を指す[18][19]。 オルトミクソウイルス科の特徴は以下の通り[20][21]。 ●エンベロープを持つ。 ●マイナス鎖の一本鎖RNAをゲノムとして持つ。ゲノムは分節性である。 ●RNA依存RNAポリメラーゼをウイルス粒子内部に含む。 ●RNAの複製が宿主細胞の核内で行われる。以前はオルトミクソウイルス科には、このA、B、C型インフルエンザの3属だけが分類されており、オルトミクソウイルス=インフルエンザウイルスとして扱われていたが、現在、トゴトウイルス属とイサウイルス属︵伝染性サケ貧血ウイルス︶という、ヒトに対する病原性が見つかっていない2属が新たにオルトミクソウイルス科に追加されているため、インフルエンザウイルスはオルトミクソウイルスのうちの一部という位置付けに当たる[21]。2016年にはD型がICTVの分類体系に追加された[18]。
型による違い[編集]
A型、B型、C型、D型の違いは、ウイルス粒子を構成するタンパク質のうち、M1蛋白︵構造タンパク質︶とNP蛋白︵核タンパク質︶の抗原性の違いに基づく。また、これ以外にも病態的、形態的、遺伝子的にも違いがあり、特にA、B型とC、D型とでは違いが大きい。型ごとの違いを以下に示す。抗原性の違い[編集]
●A型、B型、C型、D型では、M1蛋白とNP蛋白の抗原性がそれぞれ異なり交差反応しない︵例えばA型のM1やNPに対する抗体はB型、C型のものとは反応しない︶。病原的な違い[編集]
●A型、B型は毎年冬期︵まれに春期︶に流行を繰り返し、多くの場合のヒトのインフルエンザの原因になる。 ●A型は特に内部での変異型が多く世界的な大流行を起こしやすい。ウイルスに対する免疫の持続も短いと言われる。ただしA型インフルエンザウイルスに分類されるもののうち、ヒトに感染するものは少なく、残りは水鳥などの野生生物を宿主とする。 ●B型はA型に比べると流行の規模は小さいが、世界的・地域的な流行を毎年繰り返す。B型は遺伝子がかなり安定しておりウイルスに対する免疫はA型よりは長く持続すると言われる。ヒトだけを宿主とする。 ●C型は季節によらず4歳以下の小児に感染する。ほとんどの発症者(ヒト)は乳幼児期に感染するが症状が現れないことも多く、病態的にA、Bとの違いが大きいため、C型インフルエンザという別の疾患として区別して扱われることが多い。C型は遺伝子がB型以上に安定しておりほとんど変化しないので免疫は長期間に亘って持続し、一度罹ると一生持続する場合も多い。ヒトだけを宿主とする[2]。 ●D型はおもにウシで流行し、知られている例としてはブタ、ヒツジ、ヤギ、ラクダといった偶蹄類に感染し、ヒトにも感染する可能性がある[18]。形態的な違い[編集]
●C型のウイルス粒子では、電子顕微鏡下でエンベロープ上の分子であるHEが6角形に配列するのが観察される。A型、B型ではこれが認められず、A型とB型は形態上では見分けが付かない。 ●C型ではウイルス粒子の繊維状形態が特に顕著に観察される[2]。遺伝子上の違い[編集]
●A型、B型のゲノムは8分節(HA, NA, PA, PB1, PB2, M, NP, NS)、C型、D型のゲノムは7分節(HE, PA, PB1, PB2, M, NP, NS)[2][18] ●A型のNA分節にはNA一遺伝子のみがコードされているが、B型ではNAとNBの2つの遺伝子がコードされている[2]。 ●A型のM分節からはスプライシングによってM1とM2の2つのタンパクを生じるが、B型ではM1とBM2というそれぞれORFを持った2つの遺伝子がコードされており、スプライシングを起こさない[2]。亜型と株[編集]
同じA、B、C型のウイルス同士であっても、エンベロープ表面上の分子であるヘマグルチニン︵赤血球凝集素 HA:haemagglutinin︶とノイラミニダーゼ︵NA:neuraminidase︶という糖蛋白 の︵C型ではヘマグルチニン−エステラーゼ, HE︶抗原性の違いから、それぞれ複数の亜型と株に分類されている。 A型インフルエンザウイルスは特に型の内部でのHAとNAの違いが大きい。抗原性の大きな違いからこれまで16種類のHAと9種類のNAが報告されており︵1999年にスウェーデンで捕獲されたユリカモメからそれまで知られていた15種類とは異なるHAが見出され、2005年に16番目のHAとして報告された︶、その組み合わせによってH1N1〜H16N9までの亜型に分類される[22][23]。A型インフルエンザウイルスでは亜型が異なると、宿主となる生物種が異なる場合がある。B型のHAとNAおよびC型のHEは、A型に比べると多様性が低く、亜型による分類は行われない。 同じ型、同じ亜型の内部であってもHAとNAには小さな変異がある。流行を起こすウイルスには地域や年度によって違いがあり、株として分離された場所と年度によって命名・分類される。この分類によってインフルエンザウイルスのウイルス株は﹁A/ニワトリ/香港/258/97(H5N1)﹂﹁A/ワシントン/1/33(H1N1)﹂﹁B/上海/361/2002﹂のように、﹁A、B、Cいずれの属か﹂﹁分離された生物種︵ヒトの場合は省略︶﹂﹁分離された場所﹂﹁分離された順番﹂﹁分離された年度︵1999年までの場合は西暦の下2桁、2000年以降は西暦の4桁︶﹂の順に表記し、A型の場合は、最後に括弧内にHAとNAの抗原型を書く形で表される。 A型インフルエンザウイルスは、毎年流行する亜型や株が異なるが、一シーズンについて見ると流行しているウイルス︵流行株︶は、世界各地でほぼ同一であり、同時に流行しているのは数種類にとどまる。この特徴は、ワクチンによる予防を行う上でも重要であり、発生が早かった地域でのウイルス検出情報から、その年に流行する株に有効なワクチンが予測され接種されている。一方、B型インフルエンザウイルスにはこのような特徴はあまり見られず、変異の幅が少ないながら多種類の株が同時に流行する傾向がある[2][21]。A型インフルエンザウイルス[編集]
「A型インフルエンザウイルス」も参照
A型インフルエンザウイルスは、インフルエンザウイルスの中で最初に発見され、流行の規模や感染時の被害が大きいため、最も研究が進んでいる。

A型インフルエンザウイルスの構造
A型インフルエンザウイルスは、直径80-120nm程度の、エンベロープを持つマイナス鎖の一本鎖RNAウイルスである。ただし患者から分離した直後に実験室で培養したものでは1-2µm程度の繊維状の形態を示すことがあり、この場合は光学顕微鏡での観察も可能である。
インフルエンザウイルスのエンベロープは、ウイルスが放出されるときに宿主となる細胞の細胞膜を獲得したもので、その表面には10nm程度の長さの2種類のスパイクが存在しており、それぞれヘマグルチニン︵血球凝集素、HA︶、ノイラミニダーゼ︵ニューラミニダーゼ、NA︶と呼ばれる。またエンベロープ表面には少数のM2と呼ばれるエンベロープ蛋白も存在する。エンベロープの内側には、それを裏打ちする形で、M1蛋白と呼ばれるタンパク質が局在しており、これが実質的な殻の役割を果たしていると考えられている。また、最近の研究からM1蛋白の内側にごく微量の、NS2蛋白と呼ばれるタンパク質が結合していることが明らかになった。ウイルスの遺伝子は一本鎖のマイナス鎖RNAであり、8つの分節︵セグメント︶に分かれている。遺伝子はそれぞれエンベロープ内部にあるNP蛋白とよばれる核タンパク質にらせん状に巻き付いており、これがインフルエンザウイルスではヌクレオカプシドに相当する。また、それぞれのヌクレオカプシドの片端にはPA, PB1, PB2の3つのサブユニットからなるRNA依存RNAポリメラーゼが結合しており、これによってmRNAの合成やウイルス遺伝子の複製が行われる[2]。

A型インフルエンザウイルスの遺伝子
A型インフルエンザウイルスの遺伝子は8つの分節に分かれている。それぞれがコードしているタンパク質からHA︵ヘマグルチニン︶, NA︵ノイラミニダーゼ︶, PA︵RNAポリメラーゼ αサブユニット、RNA polymerase α︶, PB1︵RNAポリメラーゼ β1サブユニット、RNA polymerase β1︶, PB2︵RNAポリメラーゼ β2サブユニット、RNA polymerase β2︶, M︵マトリクス蛋白、matrix︶, NP︵核蛋白、nucleoprotein︶, NS︵非構造蛋白、non-structure︶と名付けられている。
MとNSを除く6つの分節は、名前の由来になったタンパク質1種類のみをコードしているが、MとNSの2つの分節からは選択的スプライシングによって、それぞれM1とM2、NS1とNS2の2種類のタンパク質が合成される。すなわち、A型インフルエンザウイルスが合成するタンパク質は10種類である。このうちNS1を除く9種類のタンパク質は、ウイルス粒子が構築されるときにその内部に取り込まれるが、NS1は取り込まれない︵このため非構造タンパク質と呼ばれた︶。なお、A型インフルエンザウイルスのNSは、ウイルスでは最初に見つかった、選択的スプライシングを起こす遺伝子である。
それぞれの分節において、これらのタンパク質をコードしている翻訳領域の両端には、パッケージング配列と呼ばれる独特の遺伝子配列が存在している。これらのパッケージング配列は、細胞内で新しいウイルス粒子が合成されるとき、それぞれのウイルス粒子に8つの分節がそれぞれ一つずつ正しく分配されるために必要である[2]。

インフルエンザウイルスの増殖
A型インフルエンザウイルスは、ヒトやブタでは気道上皮細胞に、トリでは大腸の上皮細胞に感染して増殖する。また実験室的には、孵化鶏卵と呼ばれる孵化途中の有精鶏卵の、漿尿液︵しょうにょうえき︶の部分にウイルスを接種して大量に培養することが可能であり、インフルエンザワクチンの製造に用いられている。また、様々な動物培養細胞に感染させる実験系も確立されている。
特に実験室的に増殖させる場合、最初は全て感染性のあるウイルスであったものが、次第に感染性を持たない不完全なウイルス粒子︵欠損粒子、DI粒子︶に置き換わっていく現象が見られることがある。これは自家干渉と呼ばれ、インフルエンザウイルス以外のウイルスにも見られる現象であるが、インフルエンザウイルスの場合は特にこれをvon Magnus現象︵フォン・マグナスげんしょう︶と呼ぶ。これは特に、高濃度のウイルスを継代していく場合によく見られる現象で、1つの細胞に複数のウイルスが感染する際、そのうちの1つが完全であれば、残りのウイルスは不完全なものであっても増殖が可能で、次第に後者が優勢になっていくためである。
A型インフルエンザウイルスの増殖過程を、以下に詳述する。

ウイルスの表面にはスパイクタンパク質と呼ばれる突起が付いており、 これが宿主細胞の表面に吸着する。
体内に侵入したウイルスは、まず標的になる宿主細胞の表面に吸着する。ウイルスは、宿主細胞に吸着するまでは、表面がタンパク質でできた単なる粒子であり、自分から宿主細胞に近づくことはできない。そのため、表面吸着の機構は非常に重要である。この過程において重要な役割をするのがヘマグルチニン (HA)およびノイラミニダーゼ︵NA︶と呼ばれる表面タンパク質である。HAとNAはウイルスのエンベロープ︵殻︶に刺さった釘のような形をしているため、スパイクタンパク質と呼ばれる[13]。HAはウイルスを構成するタンパク質の割合として最も高い40%を占め[5]、ヘマグルチニンが破壊したり変質したりすれば、そのウイルスは感染力を失う[13]。
ウイルスが細胞に侵入できるかどうかと、ウイルスが細胞内で増殖できるかどうかは、別の問題である。そのため、ウイルスのスパイクタンパク質は、自分が増殖できる細胞にのみ吸着するようにできている[13]。インフルエンザウイルスのヘマグルチニン (HA) はシアル酸に吸着する性質を持つが、間違った細胞に吸着した場合、ノイラミニダーゼ︵NA︶が吸着を断ち切って再び遊離する[5]。細胞表面の粘液にシアル酸が含まれる場合もあるので、NAの働きは、ウイルスが細胞に接触する前に粘液に吸着してしまわないようにするためにも重要である[12]。
一方、宿主細胞の表面には糖タンパク質があり、この分子の末端がシアル酸︵N-アセチルノイラミン酸︶になっている箇所がある。末端部分に付いているシアル酸なので、シアル酸残基と呼ばれる。これがウイルスのレセプター︵受容体︶の役割を果たす。シアル酸残基の隣にはガラクトースが繋がっている。このシアル酸残基とガラクトースの結合パターンはα2→6結合とα2→3結合の2種類があることが知られている。ヒトの気道上皮細胞︵つまり人の喉表面︶ではα2→6結合になっており、トリの大腸上皮細胞︵トリの大腸表面︶ではα2→3型になっている場合が多い[24]。このように、トリとヒトとでは細胞表面の構造が異なるため、トリインフルエンザが直接ヒトの細胞に吸着する可能性は低い。ただし、その可能性が皆無ではないため、養鶏場の作業員がトリインフルエンザに感染することもあり、一度感染すればその人の体内で増殖することも可能である。その場合でも、ヒトから別のヒトに感染する可能性はヒトインフルエンザに比べれば低いと考えられている[25]。ブタの気道上皮細胞には、α2→3型とα2→6型の両方の糖鎖が発現しているため、ブタにはヒトとトリ両方のウイルスが同時に感染しうる[25]。このことによって、ブタの体内ではヒトとトリ由来ウイルスの﹁合いの子﹂が生まれ、これが新型インフルエンザウイルス出現の一因になると言われる。また、ヒトの一部には遺伝的にα2→3型の糖鎖を持った人も存在することも報告されており、これが1997年以降、香港や東南アジアで発生しているトリインフルエンザのヒトへの感染の原因ではないかと考えられている。これらのヒトには直接トリ由来ウイルスが感染しうるが、大部分の︵α2→6型糖鎖を持つ︶ヒトの間での大流行には繋がらない。

ヒトインフルエンザウィルスとブタインフルエンザウイルスの遺伝的関 係性︵1918年-2009年︶
不連続変異︵抗原不連続突然変異︶は、抗原シフトとも呼ばれ、A型インフルエンザウイルスなど分節した遺伝子を持つウイルスのみに見られる突然変異の機構である。異なる亜型のウイルスが1つの細胞に同時に感染すると、細胞内で合成されたウイルス遺伝子やタンパク質が集合するときに混ざり合い、結果として元のウイルスとは異なった組み合わせの遺伝子分節を獲得した﹁合いの子﹂のウイルスが新たに生じる。例えば、H1N1とH2N2が同一細胞に感染すると、不連続変異によって理論上はH1N1, H2N2だけでなく、H1N2, H2N1という新型ウイルスが生まれることになる。
HA, NA以外のウイルス遺伝子についても同様の組み換えが起こり、結果として生じる変異が大きいため﹁ウイルスの大変異﹂とも呼ばれることがある。特に、ヒト型のウイルスと他の動物のウイルスとの間で組み換えが起きると、それまでヒトの間には存在しなかった新型のヒトインフルエンザウイルスが出現すると考えられており、実際に1957年のアジアかぜ︵H2N2亜型︶や1968年の香港かぜ︵H3N2亜型︶の出現は、この大変異によってトリ由来のウイルスがヒト型のウイルスと組み換えを起こしたことによることが、ウイルス遺伝子の研究から明らかになっている[2]。
それぞれのウイルスのレセプターの違いから、トリ由来のウイルスが直接ヒトに感染、あるいは逆にヒト由来のウイルスが直接トリに感染する機会は低いと考えられており、これまでに起きた2度の大変異がどうして起きたかについては、まだ完全に証明された訳ではない。ただし有力な仮説として、トリとヒトのウイルスの両方に感受性があるブタの体内で組み換えが起きた結果、トリ由来の遺伝子がヒト︵ブタ︶に感染する新型ウイルスを生んだのではないかと考えられている[21]。
ウイルスの構造[編集]

ウイルス遺伝子[編集]

ウイルスの増殖[編集]

ウイルスの吸着[編集]

ウイルスの侵入[編集]
ヘマグルチニンによって細胞表面に吸着したウイルス粒子は、そこから細胞内部に侵入する。インフルエンザウイルスでは、この過程は宿主細胞のエンドサイトーシスによって行われる。この過程は、宿主細胞の持つ生理機構であり、ウイルス粒子は﹁侵入﹂というよりも、いわば受動的に取り込まれる。言い換えれば、宿主細胞はウイルス粒子を積極的に取り込む[2]。なお、全てのウイルスがエンドサイトーシスを利用しているわけではなく、麻疹ウイルスやヒト免疫不全ウイルスには見られない[3]。 エンドサイトーシスにはいくつかの機構が知られているが、そのうち、クラスリン介在性エンドサイトーシスが関与することが早くから知られていた。この機構では、まずウイルス粒子が結合した部分の細胞膜は徐々に内部に向けて陥没し、それを細胞内から裏打ちするように、クラスリンと呼ばれるタンパク質が集まってくる。そして最終的に、ウイルス粒子は、細胞膜に由来する脂質二重膜と、さらにそれをクラスリンが取り囲んだクラスリン被覆小胞 (chlathrin-coated vesicle) と呼ばれる小胞に包まれた形で、細胞質に取り込まれる。細胞質内に取り込まれると、クラスリンは速やかに外れ、小胞は初期エンドソームと膜融合を起こし、ウイルスはエンドソーム内に取り込まれる。 また、クラスリン介在性エンドサイトーシスを抑制してもインフルエンザ感染が抑えられないことから、クラスリンが介在しない機構によってもウイルスの取り込みが行われることが判明している。例えば、脂質ラフトからの取り込み[26]、Rasタンパク質とPI3キナーゼ[27]などの関与が示唆されている。脱殻[編集]
前述したように、インフルエンザウイルスの表面は、エンベロープ︵殻︶で覆われており、さらには細胞に取り込まれる際にエンドソーム︵初期エンドソーム︶と呼ばれる膜に覆われる。インフルエンザウイルスにとって、エンベロープとエンドソームは細胞内に取り込まれた後はむしろ邪魔になるため、除去する必要がある。この仕組みが脱殻である。 エンドサイトーシスは本来、細胞表面の異物などを分解するための機構である。この目的のため、エンドソームの内部は弱酸性になっている。インフルエンザウイルスはこの過程から巧みに逃れるようにできている。[2] 脱殻の過程で重要な働きをするタンパク質の1つはM2タンパク質である。HAとNAがウイルスの殻の表面に刺さったような構造であるのに対し、M2タンパク質はウイルスの殻を貫通している。また、M2タンパク質は水素イオンを選択的に通過させる性質を持つ。つまり、イオンチャネル型の膜タンパク質である[13]。外側の水素イオン濃度が高い、すなわちpHが低い状態になると、M2タンパク質が開いてウイルス粒子内部に水素イオンが流れ込む。ウイルス遺伝子はNP・PA・PB1・PB2と結合してリボ核タンパク質(RNP)の状態にあり、RNPはウイルスの殻の本体であるM1タンパク質と結合をしているが、M2タンパク質の働きでウイルス粒子内部が酸性になると、RNPとM1タンパク質の結合が弱められる[3]。また、酸性になると、ウイルスの殻の主要成分であるM1タンパク質同士の結合も弱まる[25]。抗インフルエンザ薬であるアマンタジンは、このM2タンパク質のイオンチャネル作用を阻害することで、ウイルスの増殖を抑制する[28]。 脱殻の過程で重要な働きをするもう一つのタンパク質は、細胞に侵入する際にも使われたヘマグルチニン︵HA︶である。HAは宿主細胞の中で変質し、ウイルスの殻とエンドソームを結合させる糊のような役割をする[3]。具体的には、ウイルス粒子表面のヘマグルチニンは、最初HA0と呼ばれる1つのタンパク質であるが、気道や消化管の細胞が分泌するタンパク質分解酵素の働きによって切断され、HA1とHA2という2つのタンパク質になる。この現象をHAの開裂と呼ぶ。HAが開裂するとその立体構造が崩れるため、ウイルス粒子の殻が壊れやすくなり、脱殻が正常に起こるのを助ける。インフルエンザウイルスが、ヒトでは呼吸器に、トリでは消化管に感染する理由は、レセプターの発現の有無に加えて、このタンパク質分解酵素が存在するかどうかも重要であると考えられている[4]。ヒトにおいては、気道に存在するクララ細胞が分泌するトリプターゼ・クララというタンパク質分解酵素やプラスミンが、この役割を担っていると言われる。また、黄色ブドウ球菌などの細菌もHAに働きやすいタンパク質分解酵素を作り出すため、黄色ブドウ球菌などの細菌とインフルエンザウイルスの混合感染が起きると重篤化しやすい[29]。 例外として、インフルエンザウイルスの内、H5またはH7亜型ウイルスの中には、これらの特殊なタンパク質分解酵素に頼らずとも、フーリンのような多くの細胞内に普通に存在するタンパク質分解酵素によって容易にHAの開裂を起こすものがある。このようなウイルスは気道や消化管だけでなく全身の細胞で増殖できるために、急激かつ重篤な感染を起こす。強毒型あるいは高病原性インフルエンザウイルスとよばれるものには、このように変異したHAを持つものが多いことが判っており、ニワトリに大量死を発生させる高病原性トリインフルエンザがこの代表例である[29]。ヒト由来のウイルスはほぼすべて弱毒型であるが、1997年に香港で発生したH5N1亜型が高病原性︵強毒型︶であった。H5N1亜型は2011年6月までに15か国︵インドネシア、エジプト、ベトナム、中華人民共和国、タイなど︶に広まり、感染例556、死者325人が記録されている[30]。ウイルスmRNAと遺伝子の複製[編集]
細胞質に放出されたウイルス遺伝子にはNP・PA・PB1・PB2が結合してリボ核タンパク質(RNP)の状態にあるが、次にこの複合体は核内に移行し︵NPの作用と考えられている︶、ウイルスの材料であるウイルス蛋白とウイルス遺伝子の合成を始める。ただしインフルエンザウイルスはタンパク質合成に必要なmRNAを持っていないため、まずはmRNAの合成が行われる[12]。 mRNAの合成には、mRNA複製を開始するためのプライマー構造や、mRNAの終了を意味するpoly A終末が必須である。しかしながらインフルエンザウイルスの遺伝子上にはこれらが存在しない。このためインフルエンザウイルスは、PB2の働きによって、宿主細胞がDNAから作り出したmRNAを切断してプライマーとなるキャップ構造とpoly A構造を切り取り、それを自身の遺伝子に結合させてmRNAの合成を行うという、独特の方法でmRNA合成を行う。要するに、ウイルスのリボ核タンパク質は、宿主のmRNAの一部を拝借して、ウイルスmRNAを作り出す。この機構はキャップ・スナッチング (cap snatching) と呼ばれる。この方法によって合成されたmRNAは、宿主が作り出したmRNAと同様に処理されて、そこからウイルス粒子の材料になるタンパク質が大量に合成される[31]。 一方、ウイルス粒子のもう1つの﹁材料﹂となる、ウイルス遺伝子も同時に大量に複製される。この過程はmRNA合成とは異なり、ウイルス遺伝子の全長を複製する必要があるため、上とは別の機構によって、マイナス鎖RNA→プラス鎖RNA→マイナス鎖RNAという順序で合成されると考えられている。その機構についてはMCM複合体などが関与していることなどは判っているが、具体的にはまだよく判っていない[31]。 遺伝子の複製過程で、1万〜2万回に1回ほどの確率でミスが発生する。この確率はヒトの生物などと比べると非常に高く、新たな特徴を持つウイルスが生まれやすい原因となっている[12]。ウイルス蛋白合成[編集]
作られたウイルスmRNAは、宿主細胞のリボソームに張り付いて、ウイルス蛋白を作り出す[3]。大まかに分けて、初期タンパク質と後期タンパク質の2段階に分けて作られる。 ウイルス蛋白の内、核蛋白︵NP︶とポリメラーゼ︵PA、PB1、PB2︶は、宿主細胞の中で比較的初期に合成され、核内に移行する。核内に移行した後、ウイルス遺伝子と結合して新たなリボ核タンパク質(RNP)となり、再びウイルスmRNAと遺伝子の合成を始める[13]。 一方、ウイルス蛋白の内、ヘマグルチニン、ノイラミニダーゼ、M2タンパク質は、小胞体内で比較的後期に合成され、糖鎖による修飾を受けながらゴルジ体、分泌小胞を経て、細胞膜に発現する[13]。材料の集合と粒子の再構成[編集]
全ての構成材料が揃うと細胞膜の近傍で材料が集合して、ウイルス粒子の組み立てが始まる。集合部位の細胞膜からは宿主細胞自身の膜タンパク質が排除されて、代わりにウイルスのエンベロープタンパク質が集積する。また細胞質側からM1タンパクが裏打ちするように集合し、8つの分節を1つずつ含むようにリボ核タンパク質複合体が集合する。これらの集合体は、細胞膜から出芽するような形で成長していき、最終的にエンベロープで完全に覆われたウイルス粒子が再構築され、細胞外に放出される[2]。 インフルエンザウイルスの再構築の過程は、宿主細胞のタンパク質が排除されたり、8つの分節が正しく分配されることなどから、高度な分子間相互作用によって制御されていると考えられているが、その機構はまだよく判っていない。ウイルス粒子の放出[編集]
細胞外に放出された時点でインフルエンザウイルスの粒子は既に完成されているが、むしろ完成されているが故に、そのままでは他の細胞に感染することができない。ウイルスが感染した宿主細胞の表面にも、ウイルスレセプターとなる糖鎖が多く出現しているため、そのままの状態では放出されたウイルスは直ちに元の細胞表面に結合してしまい、他の細胞に感染を広げることができないからである。 そこで感染した細胞からウイルス粒子を遊離させるために働くのがノイラミニダーゼである。ノイラミニダーゼは細胞表面の糖鎖をシアル酸残基の部分で切断する活性を持つ酵素であり、この働きによって新たに作られたウイルス粒子が感染した細胞から遊離する。[2] このため、ノイラミニダーゼを阻害することは、インフルエンザの治療に有効であると考えられており、これを標的にした抗インフルエンザ薬が開発され臨床応用されている。2005年現在、ザナミビルとオセルタミビルの二種類が実用化された。2010年にはラピアクタ、イナビルが世界に先駆けて、日本で上市された。ただしノイラミニダーゼもまた変異するため、これらの薬剤に対する耐性を獲得したウイルスが出現し始めている。特に小児の場合、耐性ウイルスが発生しやすく、投与された患児の最大で16%から検出されたという報告もある[32]。また2008/2009シーズンはH1N1(ソ連)のH274Y変異株の流行により、市中H1N1感染の99.6%はタミフル耐性であった[33]。ウイルスの変異[編集]
A型インフルエンザウイルスは、ウイルスの中でも特に突然変異によって変異型ウイルスが出現しやすいものの1つである。インフルエンザウイルスが変異する場合、特に重要視されるのはヘマグルチニンとノイラミニダーゼの、2種類のスパイクタンパク質の変異である。これらのスパイクタンパク質はウイルス粒子表面にあるため、ヒトに感染したときに体内の抗体が結合して中和する標的︵抗原︶になるが、ウイルスに変異が起こると過去の感染によって作られていた抗体と反応しなくなるため、感染を起こしやすく、また重症化しやすくなる。またヘマグルチニンが大きく変異すると、レセプターとの結合性が変わった結果として、それまでヒトに感染しなかったトリや他の動物のウイルスがヒトに感染する場合もある。この他、M2タンパク質の変異によって、抗ウイルス薬の1つであるアマンタジンに対する耐性ウイルスの出現も報告されている[28]。 インフルエンザウイルスが変異を起こしやすい理由は、他のウイルスと異なり突然変異のメカニズムを2つ持っているためである。このメカニズムはそれぞれ連続変異、不連続変異と呼ばれる[2]。連続変異[編集]
連続変異︵抗原連続突然変異︶は、抗原ドリフトとも呼ばれ、ウイルス核酸が一塩基単位で変異を起こすものである。これは、一般に言う遺伝子の突然変異と同じ機構であり、インフルエンザウイルスに限らず、他のすべてのウイルスにも共通に見られる現象である。一般に、このメカニズムによる変異はDNAウイルスよりもRNAウイルスの方が出現の頻度が高い。これは、ほとんどの細胞にはDNAに異常が生じた場合の修復機構が備わっており小さな変異が修復されやすいのに対して、RNAには修復機構が存在しないためであることに因ると言われる。インフルエンザウイルスはRNAウイルスであるため、この機構による突然変異の頻度が他のRNAウイルスと同等に高い部類に属する。 連続変異によって生じる変異は、ウイルスタンパク質のどれか1つにおいて、1つのアミノ酸が変わるなどの、比較的小さな変異であるため﹁ウイルスの小変異﹂とも呼ばれることがある。A型インフルエンザウイルスでは、同じ亜型(H1N1や、H3N2など︶の内部における、変異株の違いに相当するが、変異が起きた部位がたまたまウイルスの感染性や毒性に関わる重要な部位である場合にはウイルスの性質が大きく変わる。また、小さな変異が積み重なった結果としてウイルスの抗原性が変化すると、従来のウイルスに対する抗体と反応しにくくなり、これが新型ウイルスの流行を起こすきっかけになる[2]。不連続変異[編集]

病原性[編集]
A型インフルエンザウイルスにはHAとNAの変異が特に多く、これまでHAに16種類、NAに9種類の大きな変異が見つかっており、その組み合わせの数の亜型が存在しうる。亜型の違いはH1N1 - H16N9といった略称で表現されている。ヒトのインフルエンザの原因になることが明らかになっているのは2009年現在で、﹁Aソ連型﹂として知られているH1N1、﹁A香港型﹂として知られているH3N2、H1N2、H2N2、の4種類である。この他にH9N1、高病原性トリインフルエンザとして有名になったH5N1などのいくつかの種類がヒトに感染した例が報告されているが、ヒトからヒトへの伝染性が低かったため大流行には至っていない。しかし、いずれ新型インフルエンザが定期的に大流行を起こすことは予言されつづけている。ヒトに感染しない亜型のウイルスは鳥類や他の哺乳動物を宿主にしていると考えられている。特に水鳥ではHAとNAの組み合わせがすべて見つかっており、自然宿主として重要な地位を占めていると考えられている。同じH1N1であってもさらに細かな変異によって抗原性や宿主が異なり、年によって流行するウイルスの型は異なる。 A型インフルエンザウイルスは、ヒトの呼吸器に感染してインフルエンザの原因になる。また、高病原性︵強毒性︶のトリインフルエンザウイルスがニワトリなどの家禽類に感染するとトリインフルエンザ<[注釈 1]を起こす。これらの病態や症状、治療、予防方法などについては、それぞれの項を参照のこと[2]。 ヒトやブタなど哺乳動物のインフルエンザにおいて、インフルエンザウイルスは発症した患者の気道上皮細胞で増殖する。ウイルス粒子は咳やくしゃみをしたときの唾液などの飛沫に混じって放出され、それがエアロゾルとなって、他の患者の気道に再び感染するという飛沫感染が、主な伝染の様式である。一方、鳥類のインフルエンザにおいては、ウイルスは消化管の上皮細胞で増殖し、新たに作られたウイルス粒子は糞に混じって排出される。これが乾燥して飛沫になったり、あるいは水を汚染して再びトリの体内に感染するという糞口感染がトリインフルエンザでは主な伝染経路となる。トリからブタへの種を越える感染のときもこの糞口感染が主な感染経路だと言われている。 ヒトのインフルエンザでは呼吸器症状の他に、一部の患者で合併症を起こすことがある。主な合併症は肺炎と脳炎︵インフルエンザ脳症︶である。肺炎については細菌との混合感染による場合が多いが、本ウイルスによる原発性ウイルス肺炎や続発性肺炎が起きることもある。細菌との混合感染は黄色ブドウ球菌、肺炎レンサ球菌、インフルエンザ菌による場合が多いが、特に黄色ブドウ球菌の場合はHAの開裂を促進するために重篤化しやすい[2]。 脳炎は1-5歳の乳幼児を中心に見られ致死率は20-40%に及ぶが、このとき脳神経細胞でのウイルス増殖は認められず、脳炎の起きるメカニズムはまだ判っていない。B型インフルエンザウイルス[編集]
「B型インフルエンザウイルス」も参照
B型インフルエンザウイルスは、その特徴や臨床症状の点でA型とよく似ている。特に臨床症状からはA型とB型の区別はできず、A型と同様、ヒトインフルエンザの病原体として重要である。
ウイルスの構造や増殖機構、変異についてもA型に準じるが、以下の点に違いが見られる。
構造上の特徴[編集]
B型インフルエンザウイルスの遺伝子分節のうち、NAとM分節はA型との違いが大きい。A型のNA分節が、1種類のタンパク質をコードしているのに対して、B型ではNAとNBという2種類の、翻訳開始点が異なる遺伝子がコードされていて、それぞれ合成される。またA型のM分節が選択的スプライシングによってM1とM2を合成するのに対し、B型ではM1とBM2という、翻訳開始点が異なる2つの遺伝子がM分節にコードされていてそれぞれが合成される。 BM2タンパク質はA型のM2タンパク質と構造が大きく異なる可溶性のタンパク質であり、エンベロープには発現しない。A型のM2タンパク質の役割はNBタンパク質が担っており、これはM2阻害剤であるアマンタジンによる阻害を受けない。このため、B型インフルエンザウイルスにはアマンタジンは無効である。NAはA型と同様であるため、ノイラミニダーゼ阻害剤はB型にも有効である[2]。多様性の少なさ[編集]
B型インフルエンザウイルスのHAとNAには、A型に見られるほどの多様性がない。このため亜型による分類は行われないが、HAの抗原性の違いから、それぞれの流行株はB/ビクトリア/2/87と、B/山形/16/88という2つのグループに大別することができる。A型の流行期には全世界でほぼ同一の株が流行するのに対して、B型ではこの2つのグループに属する異なる株が世界中に混在した形で流行することが多い。しかしながら、それぞれの抗原の差異はA型に比べて小さいため、B型に対する免疫やワクチンはほぼ同一、すなわちB型の中の特定の株にのみ有効なのではなくB型のいずれかに感染、あるいはワクチン接種すれば、B型全てに対してほぼ一定の効果を得られ、A型に比べて持続時間が長いことが多い。 B型はヒトには感染するが、他の動物に感染した例はほとんど報告されていない︵※︶。このため種を超えた不連続変異の問題は少なく、B型のウイルス変異では連続変異が中心だと言われてきた。このことも本ウイルスにA型ほどの多様性が見られない理由の1つだと考えられている。しかしながら異なる株が同時期同地域に共存しており、少なくともヒトの間では不連続変異による組み換えが起きていることも明らかになったため、このことと疾患との関係が明らかにされつつある[2]。 ︵※︶従来、B型の自然宿主は人のみであると考えられていたが、1999年にオランダの海岸に打ち上げられていたアザラシからB型ウイルスが分離されたという報告があり、その後の研究でもアザラシがヒト由来のインフルエンザウイルスを保有できることが示唆されている[21][35][36][37]。C型インフルエンザウイルス[編集]
「C型インフルエンザウイルス」も参照
C型インフルエンザウイルスは、構造や臨床症状の点でA型、B型との差異が大きい[38][39]。
構造上の特徴と多様性[編集]
C型インフルエンザウイルスには、A型とB型が共通して持っている、HAとNAという2種類のスパイクがなく、その代わりにHE︵ヘマグルチニン−エステラーゼ︶と呼ばれる、HAとNAの両方の役割を演じる1種類のスパイクタンパク質を有する。またM分節の発現機構が、A型B型のどちらとも異なり、選択的スプライシングによりM1とP42という2種類のタンパク質を合成した後で、P42が宿主の酵素によってM1'とCM2に切断される。このCM2タンパク質が、A型のM2と同じようにイオンチャネルとして働くと考えられている[38][39]。 C型インフルエンザウイルスのHEにもA型に見られるほどの多様性がなく亜型による分類は行われない。3-4グループが混在した形で蔓延していると言われており、このグループ間での組み換えと疾患との関係が調べられつつある。C型はB型同様にヒト以外の動物には感染しない[38][39]。C型インフルエンザ[編集]
C型インフルエンザウイルスはA型、B型とは異なり、主に4歳児以下の小児に感染して鼻汁過多を特徴とする鼻かぜ様の症状を呈する。これはC型インフルエンザと呼ばれ、A型やB型と異なり季節性がなく通年にわたって発生する。一度罹患すると免疫がほぼ一生持続し、2度罹ることは極めて稀である。小児期にほとんど全ての人が感染するが、この時期に感染しなかった場合には成人にも感染することがある。成人ではさらに咽頭痛などを伴うことがあるが、ほぼ小児のC型インフルエンザと同様である[38][39]。D型インフルエンザウイルス[編集]
「D型インフルエンザウイルス」も参照
D型インフルエンザウイルスは、C型に似た構造を持つが、ウシやブタなどの偶蹄類に感染して咳や鼻水などの呼吸器症状を呈する[18]。ヒトにも感染する可能性が示唆されているが、病原性については確認されていない[18]。
感染予防[編集]
インフルエンザウイルスは、エンベロープを持つウイルスであり、石けんや消毒用アルコールなどで、容易にエンベロープが破壊されて失活する。ウイルス感染は、空気中のエアロゾルだけでなく手や衣類に付いた飛沫からも起きることがあるため、手洗いが感染予防に有効である。特にせっけんや消毒はノロウィルスロタウィルスには効かないらしい、用いると効果が大きい[40]。 熱に対する抵抗性も病原体としては比較的低く、煮沸処理︵〜100℃︶や一般的な消毒の加熱条件︵60℃30分処理など︶、一般的な食品の加熱調理の条件︵75℃〜︶などで失活させることが可能である。このため、トリインフルエンザやブタインフルエンザに罹患したニワトリやブタなどの食肉についても、適切に加熱調理したものであれば、それらを食べることによって感染するリスクはほとんどないと考えられている。 マスクについては、ウイルス粒子そのものの侵入を完全に防御することは出来ないが、くしゃみなどによって発生する唾液の飛沫などを吸着して、その通過を妨げる効果がある。吸気の湿度を保って気道粘膜を保護することも予防につながるという意見もある。感染者のマスク宇着用は、飛沫の拡散を防ぎ、周辺との濃厚接触による伝染︵主に飛沫感染による︶の機会を減らす効果があるため、社会的な観点から有用である。非感染者の場合も、医療スタッフや患者の家族などのように濃厚接触の機会が多いケースでは、マスク着用による感染予防の効果が高い。一方、濃厚接触の機会が少ない日常の社会生活において、非感染者が感染予防を目的としてマスクを着用することについては、その有効性に対する評価は分かれている[41]。抗インフルエンザウイルス薬[編集]
詳細は「インフルエンザ」を参照
インフルエンザ迅速診断キット[編集]
インフルエンザ迅速診断キットとは、インフルエンザウイルス抗原︵主に核蛋白︶を短時間に簡便に検出するための検査であり、インフルエンザ診断補助に用いられる試薬である。
患者がインフルエンザの症状を呈している場合、医師はインフルエンザ迅速診断キットを使えばインフルエンザかどうか短時間で確認でき、陽性であれば抗インフルエンザ薬を処方できる。日本で発売が始まったのは1999年1月で[42]、このキットのおかげで、医師はインフルエンザの患者に的確な治療が施せるようになった。
インフルエンザ迅速診断キットには、いくつか種類がある[43][44][45]。
培養と実験技術[編集]
インフルエンザウイルスは、生きた細胞を使って培養する。今日では病原診断には培養細胞を、ワクチン株を作るには孵化鶏卵︵孵化途中の有精鶏卵︶を用いる場合が多い[46]。 培養細胞を使う場合には、まず培養細胞を増殖させ、トリプシンを使って細胞を液中に分散させる。その分散液の中に、インフルエンザ患者から咽頭拭い液などを使って採取した検体を入れることで、培養細胞にインフルエンザを感染させる。感染の進行は細胞を顕微鏡で見て確認する[46]。培養細胞は増殖が早く、ウイルスへの耐性がある点で孵化鶏卵よりも優れている。ただし癌細胞と似た性質を持つため、安全上の理由から、孵化鶏卵を使う方法に比べて人体に接種するワクチンへの利用は難しい。安全性を確認した上でのワクチン製造への応用が研究されており、日本では2010年1月にノバルティスファーマからMDCK細胞を使ったインフルエンザワクチンが供試されている[47]。 孵化鶏卵を使う場合には、インフルエンザ患者の咽頭拭い液などの検体を、細菌ろ過用のメンブレンフィルターを通した後、孵化鶏卵に注入する。遠心分離した検体を用いる場合もある[12]。孵化鶏卵は卵齢8〜10日のものを用いる。尖った方を下にして、上部の殻を直径1cmだけ取り除き、注射器で羊膜腔や尿膜腔に注入し、開けた穴をテープで塞ぐ。48時間ほど培養し、羊水または尿液を採取して利用する[46]。ただし高病原性ウイルスではニワトリ胎児がすぐに死んでしまい、この方法を用いることができないため大量培養は困難である。その場合、逆遺伝学の手法で毒性の弱い類似のウイルスを合成して使用する場合もある[13]。 インフルエンザウイルスは、様々な動物の赤血球と試験管内で混合すると凝集する性質がある。これは︵赤︶血球凝集反応︵HA反応, hemagglutination︶と呼ばれ、ウイルス表面のヘマグルチニンが赤血球表面の糖鎖と結合し、複数の赤血球同士を架橋させて大きな凝集体を作ることによる。この性質を利用して、ウイルスを段階稀釈したときにどこまで凝集するかを調べることで、原液に含まれていたウイルス濃度を算出できるため、インフルエンザウイルスの定量に用いられている[12]。またHA反応はヘマグルチニンに対する中和抗体によって抑制されるため、一定量のウイルスを患者血清と反応させた後でHA反応の有無を検査すれば、その患者血清中に抗体が存在するかどうかを検査することが可能である[12]。これを︵赤︶血球凝集阻止反応︵HI反応, hemagglutination-inhibition︶と呼ぶ。血清中の抗ウイルス抗体の濃度上昇は、そのウイルスによる感染が起きたことの証拠であるため、感染の有無を診断するための診断技術として用いられていた。ただし検査に時間がかかることから、迅速診断キットを使う方法に置き換えられている[5]。さらにはリアルタイムPCR法などが使われることもある[48]。インフルエンザウイルスの技術的応用[編集]
インフルエンザワクチン ︵Influenza vaccine︶[編集]
インフルエンザウイルスを人工的に培養して、インフルエンザに対するワクチン︵インフルエンザワクチン︶を作製することが可能であり、世界中でインフルエンザによる感染や重症化を予防するために利用されている。予防効果や日本における予防接種の実施などについてはインフルエンザの項を参照。 インフルエンザワクチンの作製は、孵化鶏卵︵孵化途中の有精鶏卵︶を用いて行われ、目的とするウイルス株を孵化鶏卵に接種して増殖させ、それを精製することで行われる。これはインフルエンザワクチンが初めて作られた1940年代から一貫して行われている手法である[5]。ただし、毒性が高いインフルエンザウイルスの場合、増殖中に孵化鶏卵を殺してしまう場合もあるので、その場合は逆遺伝学の手法を使って作った毒性が弱い類似のウイルスで代用することもある[13]。 一般にワクチンは、 (一)弱毒生ワクチン︵弱毒性の生きた病原体を使うもの︶、 (二)不活化全粒子ワクチン︵何らかの不活化処理をして感染性を失わせた病原体を使うもの︶、 (三)スプリットワクチン︵病原体の特定の成分を精製して使うもの。成分ワクチンと呼ぶこともある︶、 に大別される。インフルエンザワクチンではこの3種類とも実用化されているが、2010年時点で日本国内で認可され、流通しているのはスプリットワクチンのみである[49]。 弱毒生ワクチンのような生きた病原体を使う方法は、若干とはいえ感染が起こるため効果が強く、接種量が少なくて済むという利点がある。また、不活性化ワクチンがB細胞による液性免疫しか期待できないのに対し、生ワクチンはT細胞による細胞性免疫の効果も期待できる[13]。反面、インフルエンザウイルスは変異しやすいため、作られたワクチンの効果や安全性の確認に時間がかかるという欠点がある。日本の厚生労働省は副作用の懸念が強いとしてインフルエンザの生ワクチンを認可していない。アメリカ合衆国ではフルミスト[13]など認可された弱毒生ワクチンもある[49]。 不活化処理には、ホルマリンなどを用いてウイルス粒子︵ビリオン︶の構造を保持したまま不活化するもの[13]と、界面活性剤やジエチルエーテルなどでエンベロープを溶かしてビリオンを壊して不活化するもの[12]があり、前者を全粒子ワクチン︵WVワクチン, whole virusの略︶、後者をスプリットワクチン︵split virus vaccine, subvirion vaccineとも︶と呼ぶ。全粒子ワクチンは、発熱などの副作用が起こりやすいとされている[12]。スプリットワクチンはそこからさらに一部の成分のみを抽出したものであり、不活化全粒子ワクチンよりも副作用が少ないとされるが、有効性も低い[49]。遺伝子工学への応用[編集]
詳細は「タンパク質タグ」を参照
A型インフルエンザウイルスのヘマグルチニン (HA) は、早期から生化学分野で研究が進められたタンパク質である。このため遺伝子工学の分野でも早期から利用されてきた。
タンパク質の合成は、大腸菌などの生物を利用する場合がほとんどなので、できたものは目的のタンパク質とその他のタンパク質の混合物となることが多い。そこで、遺伝子工学の手法を使って、合成するタンパク質に、分離などがしやすくなるようなアミノ酸配列を組み入れて置くことが多い。それがタグペプチドである。タグペプチドは、そのタンパク質の機能そのものには大きな影響を与えず、サイズが小さいものが望まれる[50]。
この目的のため、ヘマグルチニンに含まれる、9つのアミノ酸配列︵YPYDVPDYA︶からなるペプチドをHAタグと呼んで利用する。HAタグを組み入れる手法は1988年にイリノイ大学のタイらにより報告され[51]、以後利用されてきている。HAタグが付いたタンパク質は、抗原抗体反応を利用して精製することが可能である。同様なタグペプチドとしてFLAGタグ、Mycタグ、Hisタグ、GSTタグなどが開発されているが、HAタグはこれらと並んでよく利用されているものの1つである[50]。
脚注[編集]
注釈[編集]
出典[編集]
(一)^ abトム・ウイン﹃人類対インフルエンザ﹄
(二)^ abcdefghijklmnopqrstuvwx中村喜代人 著﹁オルトミクソウイルス科﹂、吉田眞一、柳雄介 編﹃戸田新細菌学﹄︵改訂32版2刷︶南山堂、東京、2004年、784-802頁。ISBN 4-525-16012-8。
(三)^ abcdefghijk根路銘国昭﹃インフルエンザ大流行の謎﹄日本放送出版協会、2001年。ISBN 978-4140019078。
(四)^ abcde山本太郎﹃新型インフルエンザ - 世界がふるえる日﹄岩波新書、2006年。ISBN 978-4004310358。
(五)^ abcdefクロード・アヌーン﹃インフルエンザとは何か﹄白水社、1997年。ISBN 978-4560057957。
(六)^ environment.umn.edu The Big One[リンク切れ]
(七)^ 感染症情報センター インフルエンザ・パンデミックに関するQ&A
(八)^ Vijgen, Leen; Keyaerts, Els; Moës, Elien; Thoelen, Inge; Wollants, Elke; Lemey, Philippe; Vandamme, Anne-Mieke; Van Ranst, Marc (2005-02-01). “Complete Genomic Sequence of Human Coronavirus OC43: Molecular Clock Analysis Suggests a Relatively Recent Zoonotic Coronavirus Transmission Event” (英語). Journal of Virology 79 (3): 1595–1604. doi:10.1128/JVI.79.3.1595-1604.2005. ISSN 0022-538X. PMC 544107. PMID 15650185.
(九)^ Our Coronavirus Predicament Isn’t All That New(The Russian flu pandemic of 1889 might have actually been caused by a foe that has become all too familiar.) - Bloomberg Opinion(2020年5月16日)
(十)^ Yamanouchi, T.; K. Sakakami, K. & Iwashima, S. (1919). “The Infecting Agent in Influenza: an Experimental Research.” The Lancet, Volume 193, Issue 4997, Page 971.
(11)^ 山内一也﹁インフルエンザウイルスを最初に発見した日本人科学者﹂﹃科学﹄第81巻8号、岩波書店、2011年8月、807-813頁。
(12)^ abcdefghijk中島捷久、沢井仁、中島節子﹃インフルエンザ - 新型ウイルスはいかに出現するか﹄PHP新書、1998年。ISBN 978-4569559568。
(13)^ abcdefghijklmnopqr堀本研子、河岡義裕﹃インフルエンザ パンデミック﹄講談社ブルーバックス、2009年。ISBN 978-4062576475。
(14)^ Wilson et al. Structure of the haemagglutinin membrane glycoprotein of influenza virus at 3 Å resolution, Nature 289, 366 - 373 (1981)
(15)^ ﹃毒性別 新型インフルエンザ 対策完全マニュアル﹄93ページ
(16)^ 新型インフルエンザ︵A/H1N1︶の状況と季節性インフルエンザ対策への移行について(PDF:118KB)
(17)^ “コロナ対策でインフル早期終息、来季深刻化の恐れも”. ロイター (2020年4月24日). 2020年4月25日閲覧。
(18)^ abcdef村上晋, 堀本泰介﹁新しい―D型―インフルエンザウイルス﹂﹃ウイルス﹄第67巻2号、日本ウイルス学会、2017年、161-170頁。
(19)^ 堀本泰介﹁インフルエンザウイルスの種類を教えてください﹂﹃インフルエンザ﹄第20巻2号、メディカルレビュー社、2019年、88-89頁。
(20)^ ﹃戸田新細菌学﹄32版
(21)^ abcde吉田眞一、柳雄介、吉開泰信 編﹁オルトミクソウイルス科﹂﹃戸田新細菌学﹄︵改訂33版︶南山堂、東京、2007年。ISBN 978-4-525-16013-5。
(22)^ Ron A. M. Fouchier; Vincent Munster, Anders Wallensten, Theo M. Bestebroer, Sander Herfst, Derek Smith, Guus F. Rimmelzwaan, Björn Olsen, Albert D. M. E. Osterhaus (3 2005). “Characterization of a Novel Influenza A Virus Hemagglutinin Subtype (H16) Obtained from Black-Headed Gulls”. Journal of Virology (American Society for Microbiology) 79 (5): 2814-2822. doi:10.1128/JVI.79.5.2814-2822.2005. ISSN 0022-538X. PMC 548452. PMID 15709000. ISSN 1098-5514(Electronic).
(23)^ “インフルエンザウイルスについて”. これからの衛生管理. 大幸薬品. 2011年8月14日閲覧。
(24)^ 東京大学医科学研究所 鳥インフルエンザウイルスのヒトへの感染に重要なアミノ酸変異を発見 参考図2
(25)^ abc本郷誠治 新型インフルエンザ
(26)^ 大久保聡子ら 三量体Gタンパク質を介する情報伝達における脂質ラフトの役割、薬学雑誌127(1) 27-40 (2007)
(27)^ 西村尚子 インフルエンザウイルスが細胞内に侵入する新たなしくみを発見!、natureダイジェスト 2011年3月10日付
(28)^ ab辻克郎、岩橋潤、今村宜寛、吉本静志、梶原淳睦、石橋哲也、森良一、山田達夫、豊田哲也﹁A型インフルエンザウイルスに対する塩酸アマンタジン使用の問題点/Emergence of amantadine-resistant influenza A viruses﹂︵PDF︶﹃ウイルス﹄第51巻第2号、日本ウイルス学会、2001年12月、135-141頁、doi:10.2222/jsv.51.135、ISSN 0042-6857、NAID 10007862914、PMID 11977753、ONLINE ISSN 1884-3433、2011年8月14日閲覧。
(29)^ ab君塚隆太, 阿部修, 石原和幸 ほか、﹁インフルエンザウイルス感染と細菌性プロテアーゼ﹂ ﹃歯科学報﹄ 2006年 106巻2号 p.75-80、東京歯科大学学会
(30)^ 関西空港検疫所 鳥インフルエンザ(H5N1型)の流行状況 (25)、2011年6月14日付
(31)^ ab内藤忠相ら インフルエンザウイルスレプリコンと宿主複製・転写因子﹂ ﹃生化学﹄2009年80巻12号 p.1128-1133、日本生化学会
(32)^ オセルタミビル耐性に関して良くある質問、2008年1月31日付
(33)^ 2008/09インフルエンザシーズンにおけるインフルエンザ︵A/H1N1︶オセルタミビル耐性株︵H275Y︶の国内発生状況 第2報 (Vol.30 p.101-106: 2009年4月号)
(34)^ 家禽ペスト呼称の改定について 日本養鶏協会 平成15年6月13日
(35)^ Influenza B Virus in Seals(Osterhaus, A.D.M.E. et al., Science 288: 1051-1053 (May 12, 2000)。2011年8月14日閲覧
(36)^ オランダの研究者がアザラシからB型インフルエンザウイルスを発見 by Michael D. O'Neill(BioBEAT) 2011年8月14日閲覧
(37)^ 海洋哺乳類のインフルエンザウィルス感染のモニタリング カスピ海アザラシにおけるインフルエンザウィルス抗体の検出 -大石 和恵 海洋環境国際共同研究プロジェクト 国際会議﹁人間と海﹂〜沿岸環境の保全〜 ︵平成14年7月8日〜7月12日、東京・盛岡・大槌︶2011年8月14日閲覧[リンク切れ]
(38)^ abcd﹃戸田新細菌学﹄32版 787, 799-801ページ
(39)^ abcd﹃戸田新細菌学﹄33版 782-791
(40)^ 岡田晴恵 著、田代眞人 監修﹃毒性別 新型インフルエンザ対策完全マニュアル﹄ダイヤモンド社 2020年 - 第4章 感染予防対策
(41)^ 新型インフルエンザ流行時の日常生活におけるマスク使用の考え方(PDF) - 新型インフルエンザ専門家会議2008年9月22日資料
(42)^ 瀬名秀明﹃インフルエンザ21世紀﹄文春新書 2009年
(43)^ ︻日本感染症学会速報︼ インフルエンザの迅速診断キット、検体は咽頭より鼻腔から採取を 日経メディカル2002.04.13 2011年8月14日閲覧。
(44)^ インフルエンザ迅速診断キット(relenza.jpインフルエンザオンライン︶
(45)^ [医療従事者向け インフルエンザQ&A 2008版 2011年8月14日閲覧。
(46)^ abc国立感染症研究所 病原体検出マニュアル[リンク切れ]、平成15年12月9日付
(47)^ ノバルティスファーマ 新型インフルエンザワクチンに関するお知らせ
(48)^ 感染症情報センター インフルエンザの検査について[リンク切れ]、2011年3月付
(49)^ abcインフルエンザワクチンの作用メカニズムを解明 2010.04.04 大阪大学免疫学フロンティア研究センター
(50)^ ab永田恭介, 奥脇暢 編﹃目的別で選べるタンパク質発現プロトコール﹄羊土社、2010年。ISBN 978-4758101752。
(51)^ Kimple et al Overview of Affinity Tags for Protein Purification[リンク切れ], 2002
参考文献[編集]
- 中村喜代人 著「オルトミクソウイルス科」、吉田眞一、柳雄介 編『戸田新細菌学』(改訂32版2刷)南山堂、東京、2004年、784-802頁。ISBN 4-525-16012-8。
- Robert A. Lamb; Robert M. Krug (2001). “Orthomyxoviridae”. In David M. Knipe and Peter M. Howley. Fields Virology (4th ed. ed.). Philadelphia: Lippincott Williams & Wilkins. pp. 1487-1580. ISBN 0-7817-1832-5
- 河岡 義裕 (2005). インフルエンザ危機. 東京: 集英社. ISBN 4087203131
関連項目[編集]
- インフルエンザ
- トリインフルエンザ
- SARSウイルス
- ノイラミニダーゼ阻害薬
- タミフル(Tamiflu、リン酸オセルタミビル)
- リレンザ(Relenza、ザナミビル水和物ドライパウダー)
- ミスト散布 - 発生の抑止や死滅の対策として
- エンドヌクレアーゼ阻害薬
外部リンク[編集]
- 『インフルエンザウイルス』 - コトバンク
| 主要項目 |
| ||||
|---|---|---|---|---|---|
| ウイルス |
| ||||
| A型の亜型 |
| ||||
| H1N1 |
| ||||
| H5N1 |
| ||||
| H7N9 |
| ||||
| 治療 |
| ||||
| 流行・ パンデミック |
| ||||
| ヒト以外 |
| ||||
| 関連項目 |
| ||||
|
| |||||