
ティラノサウルス
| ティラノサウルス | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 ティラノサウルス・レックスの模式標本(CM 9380)の復元骨格。カーネギー自然史博物館。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 地質時代 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 中生代白亜紀末期 (マーストリヒチアン) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 分類 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 学名 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Tyrannosaurus Osborn, 1905 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| シノニム | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 種 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
ティラノサウルス︵学名‥genus Tyrannosaurus︶は、約7,000万 - 約6,600万年前︵中生代白亜紀末期マーストリヒチアン︶の北アメリカ大陸︵画像資料[注 1]︶に生息していた肉食恐竜。大型獣脚類の1属である。他にティランノサウルス、チラノサウルス、タイラノサウルスなどとも呼ばれる。
最大全長は約13メートル、最大体重は約9トンと[1]、現在まで報告されている獣脚類の中で史上最大級の体格を誇る種の一つに数えられており、中生代最後の地質区分とされるマーストリヒチアン最末期の約400万年間にかけて北米ララミディア大陸に生息していた。そしてK-Pg境界を境に絶滅している。
非常に名高く、最も有名な恐竜とされ[2][3]、﹃ジュラシック・パーク﹄等の恐竜をテーマにした各種の創作作品において頻繁に登場する[4]。一方でその生態には未解明な部分も多く、新説の多様さも相まって議論が絶えない恐竜でもある[5]。また、恐竜時代終焉の象徴として滅びの代名詞にも度々引用される。
Tyrannosaurus という名称は特に断りのない場合は属名を指す。Tyrannosaurus 属の種として広く認められているのは現在のところ Tyrannosaurus rex のみである。

アメリカ自然史博物館に展示されている頭骨
属名の Tyrannosaurus は、古代ギリシア語: τύραννος ︵テュランノス︶﹁暴君[注 2]﹂ + σαῦρος ︵サウロス︶﹁とかげ﹂ないし﹁爬虫類﹂の合成語で、﹁暴君とかげ﹂や﹁暴君竜﹂といった意味になる。本属に代表する Tyrannosaurus rex の名は一般にも広く通用している。種小名の rex はラテン語で﹁王﹂の意[6]。
恐竜を含む古生物は観葉植物や現生動物と同様、ラテン語の学名を呼称として通用するのが慣例となっており、ティラノサウルスも例外ではない。属名の﹁ティラノサウルス﹂のみ、もしくは﹁T.レックス﹂が慣用されている。
日本語の表音表記には揺らぎが多く、最も一般的な﹁ティラノサウルス﹂のほか、nの重なりに重きを置いた﹁ティランノサウルス﹂、﹁チラノサウルス﹂、﹁チランノサウルス﹂、﹁チラノザウルス﹂、﹁チランノザウルス﹂、﹁タイラノサウルス﹂、﹁テュランノサウルス﹂などがある。例として、群馬県立自然史博物館は一貫して﹁ティランノサウルス﹂表記[7][8]を採用している。呼称に関しては英語圏でも混乱があり、﹁tie−RAN−oh−saw−rus﹂﹁tye−RAN−uh−SAW−rus﹂など発音表記に差がある[9]。
また、先述のように﹁暴君竜﹂という漢訳もあり、以前ほど盛んではないものの現在も用いられている。中国語では﹁暴龍﹂︵baolong; バォロン︶あるいは﹁霸王龍﹂︵bawanglong; バワンロン︶と呼ぶ。

実化石を含むカーネギー自然史博物館のティラノサウルスの模式標本で あるが、一部に誤った復元がなされており、2003年から再構成のための作業が開始されている[10][リンク切れ]。
1892年、アメリカの古生物学者エドワード・ドリンカー・コープは後にティラノサウルスのものと同一視される脊椎の一部を発見し、マノスポンディルス・ギガス︵Manospondylus gigas︶と名付けた。1900年にはアメリカ自然史博物館の学芸員であったバーナム・ブラウンが2つ目の化石をワイオミング州で発見した。この標本はコープに師事していたヘンリー・フェアフィールド・オズボーンによって1905年にディナモサウルス・インペリオスス︵Dynamosaurus imperiosus︶と名付けられた。ブラウンは1902年にモンタナ州で3つ目の化石も発見し、同年の夏にオズボーンによりティラノサウルス・レックスとして記載された[11]。ディナモサウルスとティラノサウルスはオズボーンが1905年に発表した同じ論文の中で記載・命名された。翌1906年にオズボーンは両者が実は同種であったとしてティラノサウルス・レックスに統一したが、その際ディナモサウルスではなくティラノサウルスが有効名とされたのは、たまたま論文中で先に書かれていたのがティラノサウルスであった[12]ためである。1900年に発見された元ディナモサウルスはイギリスのロンドン自然史博物館に、1902年に発見されたティラノサウルスの模式標本は現在、米国はペンシルベニア州ピッツバーグにあるカーネギー自然史博物館にて保管されている。
1917年にオズボーンはマノスポンディルスとティラノサウルスに共通する特徴を見出し、それ以後は両者が同一視されるようになった。ただし発見されていたマノスポンディルスは一例のみで、標本はきわめて部分的であったため、オズボーン自身はそれらが同一種であると結論付けたわけではない︵後述するように、この時点でもし同一種だと認められていたならば﹁ティラノサウルス﹂の代わりに﹁マノスポンディルス﹂が有効な名前になっていたはずである︶。

フィールド自然史博物館にて展示されているスー
1990年8月12日、サウスダコタ州で非常に保存状態の良いティラノサウルスの全身骨格化石が発見された。この標本は発見者のスーザン・ヘンドリクソン︶にちなんで﹁スー﹂︵Sue︶と名付けられた。しかしスーの標本は発掘者のピーター・ラーソン博士と地主との間で所有権を巡る裁判に発展し、連邦捜査局が強引にラーソン博士の保有していたスーを押収した。その後国が一時保管した後、スーはオークションにより日本円にして約10億円という高額で落札されたことでも話題を呼んだ。現在、米国イリノイ州のシカゴ市にあるフィールド自然史博物館にて展示されている︵標本番号‥FMNH PR2081︶。
1996年、ティラノサウルス科の恐竜のものと考えられる歯の化石が日本で初めて福井県で発見された。これは白亜紀前期の地層からの発見であり、中国でも世界最古のティラノサウルス科の化石が産出していることから、ティラノサウルス科のアジア起源説も主張されている[13]。
2000年6月、米国サウスダコタ州のかつてマノスポンディルスが発見された場所から、ティラノサウルスの化石が発掘された。この化石は1892年に発見された化石と同一個体のもの︵掘り残し︶と考えられ、マノスポンディルスとティラノサウルスが同一種であることが実際に確認されることとなったが、そこでコープの命名した﹁マノスポンディルス・ギガス﹂という名前の方に優先権があるのではないかという論争が生じた。しかし、2000年1月1日に発効された国際動物命名規約第4版[14][リンク切れ]に定められた規定により、動物命名法国際審議会が強権を発動して学名 Tyrannosaurus を﹁保全名﹂としたため、名称の交代が行われることはなかった[15][リンク切れ]。
2007年4月、ノースカロライナ州立大学などの研究チームは、ティラノサウルスの骨のタンパク質を分析した結果、アミノ酸配列的にニワトリに近いという結果を得たと発表した[16][17][18]。

ティラノサウルスの近縁種であるタルボサウルスの頭骨
●恐竜類 dinosauria
●竜盤類 Saurischia
●獣脚類 Theropoda
●テタヌラ類 Tetanurae
●コエルロサウルス類 Coelurosauria
●† ティラノサウルス上科 Tyrannosauroidea
●† ティラノサウルス科 Tyrannosauridae
●† ティラノサウルス亜科 Tyrannosaurinae
●† ティラノサウルス族 Tyrannosaurini
●† ティラノサウルス属 Tyrannosaurus
以下の系統図は、Vorisらによって実行された2020年の系統解析に基づく[22]。
呼称[編集]

発見と研究の歴史[編集]


系統分類[編集]
シノニム[編集]
本種のシノニム︵異名︶を記載年の古いものから記す。左から、学術名、仮名転写、特記事項。 ●Manospondylus Cope, 1892 マノスポンディルス ●Dynamosaurus Osborn, 1905 ディナモサウルス ●?Nanotyrannus Bakker, Williams et Currie, 1988 ナノティラヌス ‥ティラノサウルス属の幼体とされているが、極めて近縁の別属である可能性が残る。[注 3] ●Dinotyrannus Olshevsky, 1995 ディノティラヌス ●Stygivenator Olshevsky, 1995 スティギヴェナトル︵スティギウェナトール︶分類学的位置付け[編集]

| エウティラノサウルス類 |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
ティラノサウルスの類縁種[編集]
ティラノサウルス属として現時点で広く認められているのは rex 種のみである。ただし、タルボサウルスをティラノサウルス・バタール︵T. bataar︶として、また、ダスプレトサウルスをティラノサウルス・トロスス︵T. torosus︶としてティラノサウルス属に含める主張もある。特にモンゴルで発見されたタルボサウルスはその大きさと形態がティラノサウルスによく似ているため、ティラノサウルスそのものではないかとも言われるが、実際にはタルボサウルスのほうが前肢の比率が小さい。古生物学関連の科学雑誌﹃アクタ・パレオントロジカ・ポロニカ﹄の記事︵外部リンク参照︶によれば、フィリップ・カリー、ジュン・フルム︵Jřrn H. Hurum︶、カロル・サバトは、系統解析をもとにタルボサウルスとティラノサウルスは別属と考えるべきであるとしている。ただし、この差異は生息していた環境の違いによるものであって両者は同属であるという説も根強く、決着は未だ付いていない。現在のところ、ダスプレトサウルスとタルボサウルスは比較的近年発見されたナノティラヌスとともにティラノサウルス亜科に分類されている。なお、ティラノサウルス科には他にアルバートサウルスやゴルゴサウルスが属している。

直立型の骨格図
直立から水平型への変遷については、恐竜#姿勢・歩行も参照。

羽に覆われた幼いティラノサウルスの想像図
体温
ティラノサウルスが鳥類のような恒温動物であったか、一般的な爬虫類と同じく変温動物であったかについて、決定的な結論は出ていないが︵恐竜恒温説も参照︶、彼らは羽毛恐竜として知られるコエルロサウルス類の一種で、鳥類とも比較的近縁であることや活動的と思われる骨格構造などから、ある程度の体温を維持できる中温性であった可能性は高い[32]。
体表
羽毛があったか否かについては1990年代中頃から議論の的となっている。ティラノサウルス上科の最も原始的な種︵ディロング︶に羽毛の痕跡が発見されていることから、少なくとも幼体には羽毛が生えていたのではないかと考えられるようになってきている[33]。こちらの説では、体の大きさで体温を保てるようになる成体は羽毛を持たないとされており、実際ワイレックスなどの研究から、成体のティラノサウルスの体表は︵少なくとも部分的には︶粒の細かい鱗で覆われていたことが判明している[34]。このように、幼少期は羽毛を持ち成長に伴って鱗肌になっていくと描写されることがあるが、この変化は化石証拠があるものではなく未だ想像の域を出ない[35]。また、かつては皮膚をなめらかにするため、或いは共喰いを防ぐために強い臭いを発する体脂を分泌していたともされていた[36][37]。
体色
ティラノサウルスの色はわかっていないが、アンドレア・カウは植生や色覚から、ティラノサウルスは暗色斑や不規則な帯を伴う暗い緑色をしていたと推測している[38]。

ティラノサウルスの頭蓋骨の成長段階を示す図
ティラノサウルスを代表とするティラノサウルス科は成体と幼体〜亜成体における身体的特徴の差異が大きかったことで知られている[32]。ティラノサウルスの場合、幼体と成体の間で1800以上の差異が見られる[21]。小型のティラノサウルス科恐竜とされていたナノティラヌスの頭蓋骨には、幼体に特徴的な線維骨構造や頭骨の未癒合などの特徴が見られ、1999年にトーマス・カーがティラノサウルスの幼体であると主張した。ナノティラヌスは華奢でナイフ状の尖った歯を有しており、頑強な体格と太い歯を持つ成体のティラノサウルスと全く形態が異なる。このことから、幼体や亜成体のティラノサウルスは成体とは異なる生態的地位に立っていたことになる[32]。全長6メートル前後の亜成体、通称﹁ジェーン﹂などは、オルニトミモサウリアに匹敵する程の俊足︵50km/h︶を誇り、アケロラプトルやパキケファロサウルスのような小型‐中型恐竜などを襲っていた可能性が高い[39]。また、時には大型のハドロサウルス科も狩りの対象になっていたとされ、とあるハドロサウルス科の化石には亜成体の歯型が明確に残されていた[40]。このことは、傍目から見れば貧弱そうな顎と歯を持つ亜成体であっても、見かけ以上の咬合力を持っていた事を示している。

ティラノサウルス科の成長曲線を示すグラフ。黒色がティラノサウルス。 Erickson et al. 2004 に基づく
7体のティラノサウルスを対象とした2004年の研究によると、ティラノサウルスの成長速度は10代で加速し、平均して1日あたり2キログラム強[41][注 5]のペースで体重が増大していた。16‐22歳に達するまでに完全な成体の体サイズに至り、それから成長が停止したと考えられている[43]。また、完全に成長した後で長生きすることはなく、同研究のティラノサウルス個体のうち3体は成長が停止した2,3年後に死亡していた。2019年2月時点で研究されたティラノサウルス個体のうち最高齢個体は28 - 29歳と推定されている[32]。2023年時点では、27‐30歳で大抵は死亡していたと考えれている[43]。
なお、著しい成長期においても常に一定のペースで成長していたわけではないことが示されている。アメリカ合衆国イリノイ州のバーピー自然史博物館が所蔵する2体の亜成体標本︵うち1体は﹁ジェーン﹂︶の骨のミネラルの偏りから、ティラノサウルスは温暖期に急成長し、寒冷期に3 - 6か月程度成長が止まっていたことが示唆されている。これは食料の少ない時期に備えた生存戦略の1つであり、ティラノサウルスの環境適応に繋がっている可能性がある[41]。

ティラノサウルスの足跡化石
ティラノサウルスの歩行・走行速度については未だ論争中である。その最大の原因は、彼らの速さを示す足跡化石が見つかっていないことにある。足跡化石そのものは発見されてはいるが、歩幅が分からないのである。加えて、走るのには不利な巨体を持ちながら、足の速い恐竜の特徴であるアークトメタターサルを併せ持っていることが挙げられる。なお、ティラノサウルスのアークトメタターサルを研究し、その論文の執筆を行ったエリック・スニベリー︵Eric Snively︶とアンソニー・ラッセル︵Anthony P. Russell︶は、ティラノサウルスがアークトメタターサルを持たない大型獣脚類と比べて遥かに機敏であることを立証しないが、ほのめかしている。このような事情があるため、下は15km/hから上は70km/hまで実に様々な走行速度説が提示されており、とりわけ約20 - 40km/hの間に収まる値が多い。以下に現在の代表的な説を紹介する。
速度15 - 20km/h説
1982年提唱。主に哺乳類における速度・歩幅・体格の関係から、足跡と比較解剖学の知見に基づいて51属の恐竜の走行速度が求められている。ティラノサウルスを含む大型の二足歩行の獣脚類は15 - 20km/hが最高速度であったとされる[44]。
速度36km/h説
1995年提唱。MOR 555︵﹁ワンケル・レックス﹂︶と呼ばれる体重6トンとされる個体を用いた研究が行われた。その結果、成体のティラノサウルスが20m/s(72km/h)で走った場合に転倒すると命に関わる大怪我をすることが示された。そのため最高10m/s(36km/h)で走っていたと推測されている[45]。また、神奈川大学の宇佐見義之は、ティラノサウルスの腰から下の筋骨格モデルを作製して仮想空間で数千万回の走行シミュレーションと衝撃耐久シミュレーションを行い、約51km/hを超えると衝撃に脚が耐えられないと判明したことから、現実的には時速36km/h程度で走ることができただろうと主張した[46]。
走行は困難であるという説
1999年提唱。ドナルド・ヘンダーソンは、アロサウルスとティラノサウルスの3D骨格モデルを作成してコンピュータ・シミュレーションを行った結果、ティラノサウルスの歩幅はそれほど広くなく、18km/h程度が限界という結果を得たと発表した。
さらに2002年、ジョン・ハッチンソン︵John Hutchinson︶とマリアノ・ガルシア︵Mariano Garcia︶は、ティラノサウルスは生体工学的に走ることができないと発表した。彼らはティラノサウルスが体重に対して後肢の関節を支えて走行するために必要な筋肉量を計算したが、その結果は走行が極めて困難であることを示しており[47][注 6]、妥当な最高速度をフルード数から求められる歩行速度限界である18km/h前後とした。なお、この研究でもモデル MOR 555 の体重を6トンと仮定している[11]。食用に改良されたニワトリをコンピューター・シミュレーションのモデルとするのに問題があるとの指摘もある。
速度40 - 50km/h説
ティラノサウルスは含気骨化した恐竜であり、鳥類と同様の気嚢を具えていたとされる。そのため、研究者によっては現在考えられているより軽量である3 - 4トンの体重を主張している。もし体重が3 - 4トンであれば、40 - 50km/hが妥当だと言われている。[要出典]近年ではマジュンガサウルスをはじめ気嚢の痕跡を残す獣脚類の骨格が複数発掘されている。恐竜の中でも、竜盤類に分類される獣脚類や竜脚類が気嚢を備えていたという仮説は多くの証拠から支持されている[48]。
速度30km/h前後説
2007年提唱。マンチェスター大学のビル・セラースはティラノサウルスの筋骨格のコンピュータ・モデルを作成し、走行のシミュレーションを行った。その結果、体重6トンのティラノサウルスは28km/hで速度で走行できるという結果を得たとした[46]︵セラースは2007年の論文発表前にシミュレーション結果をWEB上に公開している[49][注 7]︶。また、ティラノサウルス以外にも3種類の現生動物とアロサウルス、ディロフォサウルス、ヴェロキラプトル、コンプソグナトゥスの最高速度を算定したが、現生種の算定速度は実際のものと一致した。アロサウルスのモデルでも発見された足跡化石に一致する歩幅と速度が算定されている。これは現在最も中立的な説の一つであり、筋肉量、速筋・遅筋の割合、筋力などのパラメータはどれも推測される範囲の中間値を使っている。なお、2002年にハッチンソンらが発表した鈍足説と違い、筋肉の弾性要素や収縮速度及び速筋や遅筋などがモデルとして考慮されている。算定された速度は29km/hであるが、前述のようにパラメータが中間的であるため、これより速い可能性も遅い可能性もありえる。論文中には、速筋の割合や筋肉量によってどのように最高速度が変化するかのグラフが記載されており、それによると最低値で20km/h、最高値で50km/hである。
また2015年には、やはり30km/hほどの走行が可能だったとする論文が発表されている[50]。
30km/h前後という数値は、以前考えられていたよりもティラノサウルスの走行性能がカルカロドントサウルス科のような他の大型獣脚類に匹敵しうる事も示している。さらに30km/hという値は、獲物とされるエドモントサウルスを追跡するにも十分だった[50]。
長距離歩行適性説︵速度15 - 34km/h︶
2020年提唱。四肢骨の長さの比率を分析した結果、ティラノサウルスの後肢は高速疾走に向いた形態からエネルギー効率を重視した長距離歩行に適した形態へ進化する傾向があり、あまり速くはなかったという説がある。歩幅と体軸の回転性を追求した疾走型生物の場合、四肢骨は大腿部︵もも︶に対して下腿部︵脛から足先︶の方が圧倒的に長い。しかしティラノサウルスは、大腿と膝下の差が縮まりつつあった︵成体の比率は約1:1.2︶。そうした変化は大型化するに従って脛の成長が鈍化する事で発生したらしい。この事実から、ティラノサウルスは疾走型から長距離歩行型に移行していったと説明される。その推定速度は15 - 34km/hであるが、それでもトリケラトプスやトロサウルス等の角竜を追いかけるのには十分であったのではないかと論じられている[51]。また、近年[いつ?]の研究では角竜の前足は走行に適していないことが判明している。
一方でティラノサウルスは中足骨が他の大型肉食恐竜よりずっと長いため、大型恐竜の中では下腿部の比率が大きい恐竜の一つでもあり[注 8]、現生するいくつかの捕食動物よりも大きい[注 9]。なお、生物は進化の過程で大型化するにつれ異形生長するため、必ずしも四肢骨の比率変化が疾走型から長距離歩行型への移行と結び付くわけではない。
古生物学[編集]

感覚[編集]
ティラノサウルスの五感は判明している限りではどれも非常に発達していた。ウィットマーらは、視覚・聴覚・嗅覚など神経系の証拠からティラノサウルス科恐竜には獲物を素早く追うポテンシャルがあったと指摘し、目・頭・首を活用して獲物を捕らえる動物として復元している[23]。 視覚 鳥類やワニとの比較、そして視野をはじめとする頭部の再現から導き出された研究によると、ティラノサウルスは同じコエルロサウルス類のダスプレトサウルスやヴェロキラプトル、トロオドン科のように現生の猛禽類と等しい45〜60°の両眼視野を確保していた。この値は現生のワニやカルノサウルス類よりも明らかに広い[24]。視物質を3〜4種持っていたと考えられ、色を見分ける能力が高かったとされる[25]。 聴覚 脳と内耳の研究からは、ティラノサウルス科恐竜は現在のワニ類のように低周波音を聴き取ることに長けていたことが示されており、聴力はティラノサウルス科恐竜に取って並外れて重要な感覚だった[26][23]。 嗅覚 ディロングからティラノサウルスに至るまで大小様々なティラノサウルス上科の脳[注 4]を調べたところ、基盤的なティラノサウルス上科は嗅覚が比較的発達していなかった一方、派生的なティラノサウルス科は頭抜けて嗅覚が鋭かったことが分かっている[27]。また、コエルロサウルス類において比較したところ、ティラノサウルス科恐竜は原始的な鳥類やドロマエオサウルス科と同様に優れた嗅覚を持っていたことが判明しており、一方でトロオドン科や派生的な鳥類は嗅覚の面で前者3グループに劣っていた。この事は彼らの生態の違いを色濃く示唆していると言える[28]。 触覚 近縁のダスプレトサウルス・ホルネリ︵以下D. horneri︶の研究によると、ティラノサウルス科には現生のクロコダイルが顎に備えるものに似た圧力センサーがあった可能性が高い。この圧力センサーについて考えうる用途として、攻撃時の微調整や巣作り、子育てなどがD. horneriの研究では提示されている[29][30]。 味覚 他の4つと違って化石証拠の残りにくい味覚だが、タルボサウルスの脳の研究からは発達した味覚を持っていたことが示されている[31]。なお、同研究ではティラノサウルスとタルボサウルスの属差についても言及がある。体温・体表[編集]

成長[編集]


走行速度[編集]

食性[編集]
白亜紀後期マーストリヒチアン最末期の数百万年に生息したティラノサウルスは、当時の陸上生態系の頂点捕食者だった。2010年にデビッド・ホーンと渡部真人が発表した論文によると、ティラノサウルスは食事の際にも骨を噛み砕きながら肉を飲み込んでいたのではなく、肉を骨から剥ぎ取って摂食していたという。また、飲み込む際には現生鳥類やワニも採用している慣性摂食法を用い、剥ぎ取った肉を空中に放り投げて咥え直していた。2007年の研究では50キログラムの肉塊を首の筋肉を使って約4.5メートル以上投げることができたとされている[52]。腐肉食説とその議論[編集]
現在では概ね否定されているが、かつてはティラノサウルスなどの大型獣脚類は腐肉食動物︵スカベンジャー︶だと考えられていた時期もあった。古くはローレンス・ランベが1917年に提唱し、その根拠は大型獣脚類の歯があまり摩耗していなかった事にある[53][注 10]。最近[いつ?]では米国人古生物学者ジャック・ホーナーのスカベンジャー説が有名であるが、この説は一般向けの出版物やテレビ番組[注 11]などでよく取り上げられるものの、ホーナー自身一報たりと論文にしておらず、学説としてティラノサウルス腐肉専門説なるものは存在しない。 以下、ティラノサウルス腐肉食説とその反論を記す。| 腐肉食派の意見 | それに対する反論 |
|---|---|
| 獣脚類の歯に摩耗があまり確認されていない[23][53]。 | 獣脚類の歯に摩耗が確認されていないという前提は既に成立しておらず、ティラノサウルスを含め全ての大型獣脚類の歯で摩耗が確認されている。また、現生動物ではスカンベンジャーとハンターのいずれも歯に摩耗が見られる[23]。 |
| 極度に退化した前肢は、草食恐竜を襲撃するには明らかに不利である[23]。 | 前肢を使用せずとも大きな顎と歯を上手く利用すれば狩りは可能である[23]。 |
| 発達した嗅覚はハゲワシやコンドルのような腐肉食動物の特徴である[54]。 | ・新鮮な肉を探す際にこそ鋭い嗅覚が必要であった[55][56]。 ・ティラノサウルスは聴覚や視覚も卓越しており、嗅覚も感覚の1つとして駆使しながら獲物を追跡するポテンシャルがある[23]。 ・嗅覚の用途が餌の確保だけであったとは限らず、種や性別の判断に用いられた可能性がある[11][56]。 |
| 大きな体躯は他の肉食動物を獲物を取り合うのに有利だった[57]。 | ティラノサウルスの巨体および10代の頃の急激な成長期を腐肉のみで賄えるとは考えにくい。 |
| 歯の形状がハイエナのように骨を噛み砕くのに適しており、骨の中に残った骨髄を摂取することも可能であった。 | そもそもハイエナはライオンよりも優秀なハンターであり[58]、骨を噛み砕くことのできる歯はスカベンジャーに限ったものではない[55]。 |
| 当時の温暖な環境は今とは比べるまでもなく草食動物に快適でその数が多かったため、ティラノサウルスは腐肉に困らなかった。 | 化石の数から見積もられる草食恐竜と肉食恐竜の比率は、現在の自然界のものと大差は無い。 |
| ティラノサウルスは遅速で機敏さに欠けるため、素早い草食恐竜を追いかけることは不可能であった。 | 走行速度については上記の通り諸説あるが、ティラノサウルスと同時代の植物食恐竜であるトリケラトプスやエドモントサウルスは大腿部に対する下腿部の比率が小さいことから俊足の動物ではなかったと見られており、仮に走行速度が18km/h程度であっても狩りに支障はなかった[11]。 |
| 体サイズに対して目が小さく、遠方の獲物を発見できない[55]。 | 実際には絶対的なサイズも相対的なサイズも共に大きかった[56]。眼球の直径は陸上動物では最大の13センチメートル程度で、ヒトはおろか猛禽類を超える視力があった。また、他の肉食恐竜よりも両眼球の視野の重複が大きく、立体視により獲物との距離を推し量ることができた[55]。 |
さらに、ティラノサウルスが積極的に捕食を行っていたことを支持する証拠・論拠も多数存在している。
まずエドモントサウルスやトリケラトプスの化石には、ティラノサウルスに噛み付かれた後も生存し、治癒していたことを示すものが発見されている。これはティラノサウルスが生きた獲物を襲うという事実があった証拠と見なすことができる[59][60][61]。また、とあるハドロサウルス科の椎骨にはティラノサウルスの折れた歯が食い込んで発見されており、これはティラノサウルスが生きた獲物を襲って交戦した直接の証拠の1つでもある[62]。なお,捕食対象は植物食恐竜のみに留まらなかった可能性があり、本種同士が共食いをしていた痕跡を残す化石も発見されている[52]。
それと本種は、当時の食物連鎖における頂点捕食者を担っていたが、決して天下無敵の存在ではなく、Lee rexと呼ばれる個体からは、獲物のトリケラトプスの角によって反撃を受けたと思しき外傷の痕跡が報告されている[63]。
さらに本種が残した思しき巨大な糞化石︵コプロライト︶が見つかっており、内部には餌食になった植物食恐竜の骨片が大量に含まれていた。これは本種の顎の力が強大であることを示すと共に、消化にかける時間がワニのような爬虫類よりも短く、むしろ哺乳類や鳥類に近い消化器官を持っていた事を示唆している[64]。
状況証拠としては、当時の北アメリカの生態系の構成が挙げられる。当時の北アメリカに存在した恐竜のうち角竜は全体の約8割を占めていたようで、生態系のバランスを保つためには相応の捕食動物がいたはずであると推論されるが、1トン以上の体重を持ち、トリケラトプスやエドモントサウルスのような大型植物食恐竜を襲撃することのできた恐竜は今のところティラノサウルスしか発見されていない[55][注 12]。また、死亡した植物食恐竜だけを摂食するにしてはティラノサウルスの個体数が多すぎるという問題もある[55]。
これらに基づく現時点での総合的な推測としては、大多数の現生の肉食動物と同様に、腐肉があれば利用したスカベンジャーであると同時に獰猛なプレデターであったと仮定されるケースが多い。現生の肉食動物の多くが捕食と腐食の両方を行っており、それは肉食恐竜も同様であったと考えられる。こうした数多の研究から、相手が植物食性動物であれ、自らよりも小型の動物食性動物であれ、死骸であれ、ティラノサウルスや他の多くの肉食性獣脚類は、種ごとの細かな割合こそ不明なれど、概ね狩りと死体漁りの両方を行っていたと考えられている[23][55]。

ヒトと様々な恐竜との大きさ比較

ヒトと巨大な獣脚類のスケール比較。
ティラノサウルスは青色。
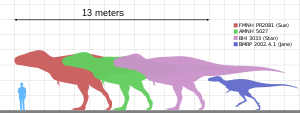
ヒトと様々なティラノサウルス標本の大きさ比較
骨格標本から推定される成体の全長は約11 - 13メートル、頭骨長は約1.5メートルで、その体重は概ね6 - 9トンと推測されている[11][1][注 13]。発見されているティラノサウルスの化石はそれほど多くはなく、2001年の時点では20体程度であり、そのうち完全なものは3体のみである。
当時の北米生態系には、ティラノサウルスに並ぶ肉食性の大型獣脚類は存在していなかった。これはジュラ紀にアロサウルス、ケラトサウルス、トルボサウルスなどが共存したのとは対照的である。また、前述のようにティラノサウルスの亜成体は非常に敏捷であり、かつ寿命における亜成体の時期がかなり長いため、本種のみが当時の生態系の中〜大型肉食恐竜のニッチを占める、一種の寡占化が起きていた[65]。

頭骨
頭部は大きく、頭骨長は胴長との比較で47%に達する[86]。頭骨は筋肉がつくと500キログラムにもなった[87]。
顎と歯
多くの獣脚類は顎の関節が外れやすく、獲物を飲み込むのに都合良くなっていたが、ティラノサウルスにはこの構造は見られない[88]。ティラノサウルスの上下の顎には鋭い歯が多数並んでいるが、他の肉食恐竜と比べると大きい上に分厚く、最大で30センチメートルにも達する。また、餌食となったとみられる恐竜の骨の多くが噛み砕かれていたことから、驚異的な咬合力[注 15]を持っていたと考えられている。推定方法によって多少の誤差は見受けられるものの、グレゴリー・エリクソンらの研究によると約8500 - 35000ニュートン[89]、カール・ベイツらの研究によると35000 - 57000ニュートンに達したとされた[52][90]。この数値は現生の大型クロコダイルや他の獣脚類︵ギガノトサウルスなど︶を軽く凌駕していた。このような強力な咬合力では頭骨に莫大な負荷が掛かるが、ティラノサウルスの頭骨は40以上のパーツに分かれた上にそれぞれのパーツに数ミリメートルほどの間隙があり、この仕組みで負荷を分散させている[90]。エミリー・レイフィールドらによる有限要素解析法(EFA)を用いた力学的研究では、発生する応力に頭骨が耐えられることが示された[91][32]。これらのことから、ティラノサウルスは獲物に対し、他の肉食恐竜のように獲物の皮膚を切り裂いて出血死を狙う方法は用いず、短時間で仕留めていたと考えられている[52]。

ティラノサウルスの歯
ティラノサウルスは各部位によって僅かながら歯の分化が進んでいたとされる[92][93]。特に門歯は断面が特徴的なD字型をしており、ティラノサウルス類を見分ける上での指標になっている。前上顎歯数は4、上顎歯はティラノサウルス類ではティラノサウルス・レックスが最も少なく11本。下顎歯もT.rexが最少の11本である。
生きている間、歯は何度でも生え変わった[92][94]。
William L. Ablerの行ったティラノサウルスの歯で肉を切る実験では、鋸歯部分に肉のカスが引っかかる結果となっており、腐敗性細菌が繁殖することで、ティラノサウルスの噛みつきを受けた獲物に致命的な感染症を負わせたかもしれない[95]。
下顎の神経血管系は非常に複雑で、優れた触覚感度を持つワニに匹敵する[96][97][98]。
舌骨は短く、ワニ類と類似していることから舌は口底部に固定されていた可能性がある[99]。
頭蓋と脳
頭蓋は同じ大きさの他の獣脚類に比べて明らかに幅広であり、特に後眼窩部の張り出しが著しい。吻部も丸みを帯びた広い形になっている。背側側頭窓には血管と脂肪を含んだ組織があり、温度を調節し脳を守るのに役立ったと考えられている[100][101]。推定される脳の重量は424gで、既知の恐竜で最大の脳を持つ[102]。脳の神経細胞の数はサルの仲間に匹敵していたとする説がある[103][104]が、この数値は再検討では10分の1程度とされた[105]。
眼
眼窩は他のどのティラノサウルス類よりも正面を向いている[106]。鍵穴型の眼窩を持ち目は小さかった。この鍵穴型の眼窩は、咬合時に眼窩後の硬い部分へと圧力を分散させ頭蓋骨の受ける負荷を軽減する効果があった[107][108][109]。眼球は鍵状の眼窩のうち、上側の窪みに収められていた[110]。現生陸生脊椎動物の目の形態の傾向を適用すると、ティラノサウルスは円形の瞳孔を持っていたと考えられる[111]。視野などは﹁視覚﹂の節を参照。
軟組織
頭蓋骨に対する歯の長さや神経血管孔の形態を現生のワニやトカゲと比較した研究では、トカゲのものに似た唇が存在していた可能性が高いと結論づけられている[112][113][114][115][116][117]。鼻から目にかけて、頭部の上部に角質の覆いがあったと考えられている[118]。ケネス・カーペンターは著書において、ティラノサウルスには喉袋があり、摂餌や求愛に役立ったとしている[119][120]が、喉袋と解釈される構造のある標本は一般的にはタルボサウルス属とされる種のものである[121][122]。

T.mcraeensisの骨格図

T.mcraeensisの復元図
2024年には、ニック・ロングリッチらによってT.mcraeensisが記載された[184][185]。rexより古い時代から得られたもので、大きさは同程度とされる[185][186]。T.mcraeensisはT.rexと比べて顎が細く、眼窩後の隆起が顕著でないとされる[187][188]。記載時点で最もT.rexに近縁な種だとされるが、rexの直接の祖先である可能性は低い[185]。
体格[編集]



病理[編集]
病気 多くのティラノサウルスの顎の化石には無数の病変の痕跡が残されている。当初この病変は別のティラノサウルスの個体に襲われた負傷だと考えられていたが、﹁スー﹂の下顎の骨に開いた穴の研究から、トリコモナスに類縁の種が影響したと2009年に発表された。トリコモナス・ガナリエはニワトリ、ハト、シチメンチョウ、ハヤブサなどの多くの現生鳥類の顎に見られる寄生性原虫であり、寄生すると宿主の顎の骨を消費して変形や損傷をもたらし、餌の嚥下や呼吸までもを困難にする。﹁スー﹂の場合も同じような症状で餓死した可能性を研究チームは示唆している。同様の生態の生物にティラノサウルスが苦しめられていたことが示唆されている。また、この感染症の痕跡が同じティラノサウルス科のアルバートサウルスとダスプレトサウルスの化石からも発見されており、トリコモナスによる感染症がかなり昔から存在していたと考えられるとともに、鳥類が恐竜から進化したという可能性をさらに強める証拠となった[23][66][67][68][69][70]。2022年には、トリコモナスの痕と解釈された穴は感染症ではなく外傷とみなすのが妥当ではないかという指摘がされた[71]。 外傷 スタンやペックズ・レックスなどの標本には同じティラノサウルスに噛まれたと見られる傷が見られる[72][73]。ワイレックスは尾が欠損しているが、これは共食いによるものの可能性がある[74]。性差[編集]
かつては尻尾の付け根にある血道弓と呼ばれる骨の位置からオス・メスを判断できると考えられ[75][注 14]、例えば﹁スー﹂はメスだと判断されてきた。しかし、今では根拠とされていた血道弓の配置が全く異なることが判明し、﹁スー﹂をメスと断定する根拠は失われてしまった[76]。 骨盤で見分けることができるという説があり、その説ではスタンは骨盤が狭いことから雄であるとされる[77]。 雌個体の方が全体的な体型ががっしりしているとする説があり、具体的にはバッキーと呼ばれる標本などが該当すると主張する研究者もいるが、万人に受け入れられている説とは言い難い[78]。 Bレックスと呼ばれる標本の大腿骨から骨髄骨が発見されており、現生鳥類では産卵を控えた雌に見られる特徴であることからBレックスは雌個体だったと考えられている[79][80][81][82][83][84][85]。形態[編集]
頭部[編集]


胴体[編集]
胴体は頑丈なつくりをしている[123]。 脊椎骨数は、頚椎‥10、胴椎‥13︵ただし、第13胴椎は僅かに仙椎的に変化している︶、仙椎‥5、尾椎‥35-44。頚椎、胴椎といくつかの仙椎は側腹腔︵pleurocoel︶を生じ、椎体にはcamellate構造がある。つまり、含気化が進んでいるのである。 首は短いが柔軟で、獲物の肉を捻り切ることに適している[124]。頸椎の棘突起は頑丈で、靭帯の付着点であった[125]。 肋骨は長大で、首を動かす筋肉が付着していた[125]。腹部には腹肋骨が存在する[36][125]。 他のテタヌラ類同様、恥骨の遠位部が前後に長く伸びるが︵ピュービックブーツ(pubic boot)と呼ばれる︶、ティラノサウルスの仲間はこれが特に巨大である。 文献によっては皮骨板があったとしているものもある[36]が、これはディナモサウルスとして記載された標本にアンキロサウルスのものと思われる皮骨板が混入していたことに由来する[126]。四肢[編集]
前肢 体の大きさに比して前肢は異常に小さく︵長さは人の腕と同程度[127]︶、用途は未だにはっきりとしていない。ただし、その大きさのわりにはかなり大きな力を出せたことがわかってきている。逆に頭部は非常に大きく、それを前肢の代わりに上手く活かしていたのではないかと考えられている。また、進化の過程で体の前方が重くなったため、前肢を短く軽くすることでバランスを取ったとする見解もある[128]。近年では前肢の用途として攻撃に用いた[129]、短い理由として集団で死骸を漁る際に噛まれない為[130]などの説が出ているが、未だ結論は出ていない。 指は2本。幼体には第3指が存在するが、成長に伴って第3中手骨と癒合する[118]。第3指は、付け根が前肢全体を横向きに回転させる筋肉の付着点となっていた[131]。この前肢は、小さいながらものを掴むことのできる構造であった[132]。 後肢 体重を支えるために太く頑丈なつくりをしていた[125]。指は4本で、前向きに3本、後ろ向きに1本生えている[125]。ティラノサウルスとその類縁種︵ティラノサウルス上科︶は、足の速いオルニトミモサウルス類[注 16]と共通の特徴であるアークトメタターサルを有していた。アークトメタターサルとは、第三中足骨が、第二・第四中足骨によって挟み込まれ、上端が押し潰される形態のことを指す[11][注 17]。近年の研究[いつ?]によると、第三指骨および中足骨に負荷が加わると靭帯の働きにより第二、第四中足骨が中央にまとめられ、負荷の方向を一直線にすることで俊敏性を増すのに役立っていたと考えられている。また、靭帯の損傷も防げたのではないかと推測される。このアークトメタターサルはオルニトミモサウルス類との共通先祖から受け継いだ形質と思われていたが、それを持たないティラノサウルスの先祖種の発見から、現在では収斂進化によるものとされている。 最重量級のティラノサウルスとされる﹁スコッティ﹂の推定体重は、大腿骨の周囲長から推定されている[注 18]。周囲長は﹁スコッティ﹂が590ミリメートル、﹁スー﹂が580ミリメートル、﹁スタン﹂が505ミリメートルである[47]。 この後肢を用いて、現生の猛禽類のように獲物を掴んだり引き裂いたりした[133]、あるいは獲物を蹴転がした[37][134]など、狩りの際積極的に攻撃に用いたとする意見も存在する。尾椎[編集]
尻尾はテタヌラ類に共通した、内部の骨が絡み合った細長い構造をしていた。そのため、より基盤的な獣脚類であるケラトサウルスよりも柔軟性は失われていたが、それでも撓るような動きは可能であった[76]。 ティラノサウルスは恐竜の中でも特に尾大腿筋肉が発達していた。尾大腿筋肉とは大腿骨から尻尾の付け根にかけて伸びる太い筋肉であり、ティラノサウルスが走る際の主電源、言わばエンジンのような役割を果たしていたと考えられている。そのためアロサウルスのような他の大型獣脚類に比べ、ティラノサウルスは強い脚力を持っていたと考えられている[135]。生態[編集]
先に述べておくが古生物の行動は実物を観察できないため、どうしても不確定な部分が発生する。そのため生態の項の多くは可能性の範疇であることを忘れてはならない。狩り[編集]
ティラノサウルスの主要な獲物の一つであるトリケラトプスを襲うにあたり、もっとも合理的なのは相手の側面を攻撃することであるとする意見がある[136][137]。事実、トリケラトプスの角は仲間内でさえ骨に少なからず傷を残す威力であり[138][139][140][141] 、これを避けるためには側面を突くのが手っ取り早い。また、側面攻撃の利点は、その手早さだけでなく、短い前腕をも効果的に︵補助的な役割で︶使えたことにある。 群れを作って狩りをしていた可能性がある︵生態>社会性も参照︶[142]が、本属を含め獣脚類が群れを作って狩りをした明確な証拠はなく、あくまで仮説である点に注意が必要である[143]。遊び[編集]
絶滅動物としては非常に珍しいことに、ティラノサウルスは時たま遊んでいた可能性が指摘されている。その根拠として、捕食されたと思われるケラトプス科の骨に不自然な噛み傷が多数残されていたからである。絶滅動物である以上ティラノサウルスが本当に遊んでいたのかは判断がつかないところではあるが、ティラノサウルスのような上位の捕食者には少なからず時間的な余裕があることや、ティラノサウルスの脳が大きいこと、現生の鳥類やワニが複雑な行動をする事から考えると、ティラノサウルスも日々の生活において何らかの娯楽を求めていたのかもしれない[144][145] 。食性(嗜好)[編集]
噛み跡の偏りから、ティラノサウルスがトリケラトプスの首の背側の筋肉を好んで食べていた可能性が指摘されている[146]。その発達した頭部を支えるため、トリケラトプスの首には強健な筋肉を蓄えており、この﹁ネック﹂と呼ばれる部位は噛みごたえと味の濃さが魅力で、これが正しいとするなら、ティラノサウルスはかなりのグルメだったようだ[147]。また、1990年にアメリカで発掘された化石から骨が丸く溶けている箇所が確認されたことから、痛風を患っていたとみられているが、これは赤身肉と内臓肉を多く食していたためと考えられている[148][149]。水泳[編集]
体の大きさを有利に使うために浅瀬で狩りを行ったとする仮説に基づき生体力学モデルを用いて行われた推定では、成体のティラノサウルスは水中では小型の獲物より速く動くことができたとされている[150]。社会性[編集]
ティラノサウルスは以前はトラなどの現生肉食動物の様に単独で行動していたと考えられていた。しかし近年では、家族または同種族の様々な世代で集団を構成し、社会生活を営んでいたのではないかとする意見もある[23][151]。この説は、アルバートサウルスなど大型獣脚類でも集団化石が見つかっていることや、後ろ脚の骨の歩行困難と思われるほどの骨折が治癒した形跡を示す個体が発見され、狩りができない期間に仲間が餌を運んでいた可能性があることに基づく推論である。DINO A LIVEなどではこの説に基づきティラノサウルスの親離れが描かれている。 ブラックヒルズ地質学研究所はスタン標本が生前、家族と共に過ごしていた可能性を示唆している[152]。雌が雄よりも大きかったとする説に基づいて、家族内の雌が群れのリーダーであったとする説もある[153]。 また、亜成体の頭蓋骨に残された﹁捕食や殺傷を目的としない攻撃によってつけられたと考えられる治癒した傷﹂の痕跡から、亜成体は成長段階の近い個体からなる階層を持つグループを形成しており、上位のものが下位のものに対して致死的ではない噛みつきを行うことで階層が維持されていたとする説もある[154][155]。産卵と子育て[編集]
硬い卵を産む現生鳥類同様に骨髄骨が見られるが、ティラノサウルスの卵が硬かったのか柔らかかったのかは未だ不明である[156]。 ティラノサウルス属を含むティラノサウルス類4種の生存曲線から、鳥のように熱心に子育てをするのではなく爬虫類のような様式であったと考えられている[157]。 田中康平は、植物のマウントを作り発酵熱で卵を温めたという説を唱えている[156]。 ティラノサウルスは下顎に無数の神経が走っていることがわかっており︵形態>頭部>顎と歯も参照︶、感覚が鋭敏だったと推測されることから、子供に顎で触れてコミュニケーションをとったり顎で子供を運搬したりした可能性も示唆されている[98][158][159][160]。親子による狩り説[編集]
この説はフィリップ・カリーがティラノサウルスの生体的特徴や近縁種の集団化石から推測した話を元に、ティラノサウルスが親子で狩りをしていたとする説がある[161]。なお、カリーの推測は、トーマス・ホルツの書いた﹃ホルツ博士の最新恐竜事典﹄でも確認でき[162]、とりわけNHKが恐竜特番を組む際に採用することが多い[163][164]。他にはディスカバリーチャンネルの﹃恐竜再生﹄でも取り上げられている。 この説は前述の鈍足説に基づく。ティラノサウルスの咬合力は非常に強力であるが、成体のティラノサウルスは鈍足であると考えられるため、簡単には獲物に接近できない。そこで、小型かつ軽量なため機敏な動作ができる子供が獲物を親の元まで追い立てたところで親が仕留めていたのではないか、との説明がなされている。 この仮説の問題点としては以下のような指摘がある。 ●実際には獲物の大型植物食恐竜のほうが鈍足だったこと。なお、エドモントサウルスについては時速30km前後と成体のティラノサウルスと互角の敏捷性があったらしい[165]。 ●体重4-5トンを軽く超える大型恐竜を脅す役割を体重1トン程度の亜成体が担っていること[注 19]。 ●群れに子供がいて初めて成り立つ狩猟方法であること。その他の可能性[編集]
●狂乱索餌 大量の獲物︵もしくは死体︶によって一帯の肉食動物が自然と集結し、貪るように獲物へ向かっていく行動︵詳しくは狂乱索餌を参照︶。これに近い行動ならば、群れの存在と集団での狩りを説明できるかもしれない[要出典]。化石の評価[編集]
1997年10月4日には、シカゴのフィールド自然史博物館に展示されているスーと呼称される個体が当時836万ドル︵2020年現在の価値では1350万ドル、日本円で約14億円︶もの高値で落札され、化石史上最も高額とされていた。2020年10月6日にはブラックヒルズ地質学研究所のスタンが3180万ドル︵日本円で約33億円︶で落札され、最高額を更新した[166]。 2012年5月、アメリカのオークションにティラノサウルスの骨格化石が出品され105万ドルで落札されている。ただし、この標本はモンゴルから密輸されたことが明らかになり、後日差し押さえを受けている︵この化石はティラノサウルスではなく、タルボサウルスとされることもある種のものである。記事ではティラノサウルスと紹介している︶[167][168][169]。 このようにティラノサウルスは商業的取引が盛んであるが、販売は科学の手を離れることを意味し批判も多い。トーマス・カーによれば、2020年10月時点で研究に利用可能な信頼できる標本が57体、商業的・私的に保存されている標本が43体であるという[170][171]。T.rex のほかのティラノサウルス[編集]
これまでにティラノサウルスはT.rex一種しか認められていなかった。 2022年、アメリカのグレゴリー・ポール氏による37体の化石の分析によると、歯と大腿骨の違いなどから、T.regina(女王という意味)とT.imperator(皇帝という意味)の2種類を発見したと発表し、命名した[172][173][174][175][176][177] [注 20]。 しかし、これくらいの違いは一つの種の中でも見ることができるという意見もある。カーセッジ大学の研究チームは、差は小さく、これらの種小名は無効でティラノサウルスにはT.rex一種のみだとした[179][180][181][182]。イタリアの古脊椎動物学者アンドレア・カウも自身のブログにおいて、骨が堅牢か虚弱か、切歯状の歯が1本か2本かの2つの解剖学的特徴のみに基づいていることを述べ、堅牢・虚弱の明確な区別がなく、恐竜の歯は成長によって変動しやすい点から種を区別するのに十分でないと指摘した[183]。

大衆文化におけるティラノサウルス[編集]
ティラノサウルスを主題とする作品一覧も参照。 2020年代には米国発祥とされる[189][190][191][192]、﹁ティラノサウルスの着ぐるみ着用の徒競走大会﹂︵ティラノサウルスレース︶が、日本各地でブームとなった[190][191][193][194][195][196][197][198][199]。脚注[編集]
注釈[編集]
(一)^ 約6,500万年前︵K-T境界上︶の大陸配置図︵K-T (65Ma)︶。この時までの一時期をティラノサウルスは左上の大陸で生きていた。- Mollewide Plate Tectonic Maps - Dr. Ron Blakey[リンク切れ]
(二)^ ﹁暴君﹂は近代的な語義。本来のギリシア語の意味については僭主を参照のこと。
(三)^ ニック・ロングリッチは2024年にナノティラヌスはティラノサウルスと別属の成体であるとした[19][20]が、複数の研究者が反論している[21]。
(四)^ 正確には型取りしたエンドキャスト。
(五)^ 2011年には平均して1日あたり5キログラムのペースで増量したとする研究も発表された[42]。
(六)^ 走行運動は脚の垂直方向に大きな荷重をかけ、人間の場合は体重の2.5倍ほどの荷重が立脚相中期にかかるとされている。もし、体重6トンのティラノサウルスの脚にその2.5倍の荷重がかかるとすると、左右の脚にそれぞれ体重の約40%[47]、合計で86%ほどの筋肉が必要になると論文中には述べられている。筋肉量は各関節にかかるトルクと関節から推定されるモーメントアームから計算されている。必要となる片足の筋肉の内訳は、股関節伸展筋が体重の15%、膝関節伸展筋が4%、足首関節伸展筋が15%、屈指筋が9%であり、合計43%必要であると算定されている。そのような筋肉量はありえないので、ティラノサウルスは走れないと結論付けられた。ただし、現生のダチョウのような大型陸鳥では筋肉と腱などの連動性が下肢の筋肉量を小さく抑えるのに役立っているが、ティラノサウルスの筋肉の弾性要素と腱の連動性については不確定要素が多かったため、上のハッチンソンの計算では無視されている。
(七)^ 簡単なティラノサウルスの筋骨格モデルを作成した結果、体重6トンのティラノサウルスは最高25 - 54km/hで走れると書かれている。ムービーで公開されているモデルは38.5km/h︵10.7m/s︶である。
(八)^ アロサウルスなどを含むカルノサウルス類と比べると明らかに比率は上回っている。
(九)^ ライオンを上回り、ウマよりやや劣る。
(十)^ なお説の発端はティラノサウルスではなく近縁のゴルゴサウルスである。
(11)^ ﹃新説・T-レックス﹄︵ディスカバリーチャンネル制作︶など。
(12)^ ダコタラプトルは鉤爪こそ大きいものの、体格は華奢であり、そもそも全長5メートル弱であった。
(13)^ 体重に関しては異説も多く最低3トンから最大14トンまで幅がある。
(14)^ メスは卵を通すために血道弓が短いと考えられていた。
(15)^ 上下の顎の咬み合わせの力をいう。
(16)^ ダチョウに似た恐竜群。
(17)^ メタターサル︿metatarsal﹀は中足骨のこと。
(18)^ これはティラノサウルスに限った話ではなく、二足歩行の動物であればこの手法が通用する。
(19)^ ただし亜成体の咬合力が従来の予想を上回るものであった可能性も指摘されている。上記﹁成長﹂の節を参照。
(20)^ 属名と種小名両方の文法形式は、男性形、女性形、中性形のいずれかで統一される必要があるが、Tyrannosaurusが男性形なのに対してreginaは女性形であり適切ではない[178]
出典[編集]
(一)^ ab“史上最大のティラノサウルスと判明、約9トン”. ナショナル・ジオグラフィック協会 (2019年3月28日). 2020年7月20日閲覧。
(二)^ Bakker, Robert (2000年). "Prologue". In Fiffer S (ed.). Tyrannosaurus Sue. New York: W. H. Freeman & Company. pp. xi–xiv. ISBN 0-7167-4017-6。
(三)^ Lowell Dingus and Mark Norell, Barnum Brown: The Man who discovered Tyrannosaurs rex, (Los Angeles: University of California Press, 2010, pg 94)
(四)^ Brian Switek (2013年4月10日). “T・レックス、映画と実際の姿の違い”. ナショナルジオグラフィック. ナショナルジオグラフィック協会. 2021年4月29日閲覧。
(五)^ ﹁T・レックス、羽毛ではなくうろこに覆われていた 最新研究﹂AFPBB news、2017年6月7日。2020年8月22日閲覧。
(六)^ Osborn, H. F (1905年). "Tyrannosaurus and other Cretaceous carnivorous dinosaurs". Bulletin of the AMNH. 21(14): 259–265. (七)^ “ティランノサウルス・レックス︵動刻︶”. 群馬県立自然史博物館. 2020年12月19日閲覧。
(八)^ 土屋健︵著︶﹃リアルサイズ古生物図鑑 中生代編﹄群馬県立自然史博物館︵監修︶、技術評論社、2019年8月3日、248–251頁。ISBN 978-4-297-10656-0。
(九)^ ヒサクニヒコ﹃新・恐竜論 地球の忘れものを理解する本﹄PHP研究所、2004年2月23日、50頁。ISBN 978-4-569-63334-3。
(十)^ Rebuilding T .Rex - カーネギー自然史博物館のサイトのトピック。
(11)^ abcdef真鍋真、樽創﹁Tyrannosaurus rex 発見百周年﹂﹃化石﹄第72巻、日本古生物学会、2002年9月20日、45–47頁。doi:10.14825/kaseki.72.0_45。2020年12月19日閲覧。
(12)^ Osborn, H.F (1906年). "Tyrannosaurus, Upper Cretaceous carnivorous dinosaur (second communication)". Bulletin of the AMNH. 22: 281.
(13)^ ﹁<中国暴竜>ティラノサウルス類の起源は東アジア、北米移住説覆す―地質科学院﹂﹃Record China﹄2009年10月6日。2020年7月19日閲覧。
(14)^ 動物命名法解説
(15)^ ICZN : Palaeontology & Biostratigraphy
(16)^ “鳥と恐竜は近縁、6800万年前の化石から証拠発見 - 米国”. www.afpbb.com (2007年4月13日). 2023年10月27日閲覧。
(17)^ “利用案内・情報 ≫ ホットニュース ≫ 2009-05-15 :: 国立科学博物館 National Museum of Nature and Science,Tokyo”. www.kahaku.go.jp. 2023年10月27日閲覧。
(18)^ “鳥と恐竜を結び付けたティラノサウルスのタンパク質”. natgeo.nikkeibp.co.jp. 2023年10月27日閲覧。
(19)^ Bath, University of. “New research shows 'juvenile' T. rex fossils are a distinct species of small tyrannosaur” (英語). phys.org. 2024年1月4日閲覧。
(20)^ Longrich, Nicholas R.; Saitta, Evan T. (2024年3月). "Taxonomic Status of Nanotyrannus lancensis (Dinosauria: Tyrannosauroidea)—A Distinct Taxon of Small-Bodied Tyrannosaur". Fossil Studies (英語). 2(1): 1–65. doi:10.3390/fossils2010001. ISSN 2813-6284。
(21)^ abpublished, Sascha Pare (2024年1月3日). “Nanotyrannus vs. T. rex saga continues: Controversial study 'doesn't settle the question at all'” (英語). livescience.com. 2024年1月4日閲覧。
(22)^ Voris, Jared T.; Therrien, Francois; Zelenitzky, Darla K.; Brown, Caleb M. (2020年). "A new tyrannosaurine (Theropoda:Tyrannosauridae) from the Campanian Foremost Formation of Alberta, Canada, provides insight into the evolution and biogeography of tyrannosaurids". Cretaceous Research. 110: 104388. doi:10.1016/j.cretres.2020.104388. S2CID 213838772。
(23)^ abcdefghijデイヴィッド・E・ファストヴスキー [ドイツ語版]、デイヴィッド・B・ウェイシャンペル [英語版]︵著︶﹃恐竜学入門 ─かたち・生態・絶滅─﹄藤原慎一・松本涼子︵訳︶、真鍋真監訳︵第2版版︶、東京都文京区千石3丁目36-7: 東京化学同人、2018年5月25日︵原著2015年1月31日︶、185–190頁。ISBN 978-4-8079-0856-1。NCID BB17901360。OCLC 904989280。全国書誌番号:22535495、ASIN 4807908561。
(24)^ Kent A Stevens (2006年). "Binocular vision in theropod dinosaurs". Journal of Vertebrate Paleontology. 26(2). doi:10.1671/0272-4634(2006)26[321:BVITD]2.0.CO;2。
(25)^ https://www.edu.city.kyoto.jp/science/about/0212tyranno.pdf
(26)^ Lawrence M. Witmer; Ryan C. Ridgely (2009年8月26日). "New Insights Into the Brain, Braincase, and Ear Region of Tyrannosaurs (Dinosauria, Theropoda), with Implications for Sensory Organization and Behavior". The Anatomical Record. 292 (9). doi:10.1002/ar.20983。
(27)^ Martin Kundrát; Xing Xu; Martina Hančová; Andrej Gajdoš; Yu Guo; Defeng Chen (2018年). "Evolutionary disparity in the endoneurocranial configuration between small and gigantic tyrannosauroids". Historical Biology. 32(5): 620–634. doi:10.1080/08912963.2018.1518442。
(28)^ Darla K Zelenitsky; François Therrien; Yoshitsugu Kobayashi (2008年10月28日). "Olfactory acuity in theropods: palaeobiological and evolutionary implications". Proceedings of the Royal Society B. 276 (1657). doi:10.1098/rspb.2008.1075。
(29)^ Thomas D. Carr; David J Varricchio; Jayc C Sedlmayr; Eric M Roberts; Jason R Moore (2017年). "A new tyrannosaur with evidence for anagenesis and crocodile-like facial sensory system". Scientific Reports. 7. doi:10.1038/srep44942。
(30)^ “ティラノサウルスはこんな顔だった、最新報告”. ナショナル・ジオグラフィック協会 (2017年4月4日). 2020年7月20日閲覧。
(31)^ S.V. Saveliev; V. R. Alifanov (2007年). "A new study of the brain of the predatory dinosaur Tarbosaurus bataar (Theropoda, Tyrannosauridae". Paleontological Journal. 41: 281–289. doi:10.1134/S0031030107030070。
(32)^ abcdeダレン・ナイシュ、ポール・バレット︵著︶﹃恐竜の教科書 最新研究で読み解く進化の謎﹄吉田三知世︵訳︶、小林快次・久保田克博・千葉謙太郎・田中康平監訳︵第1版版︶、大阪府大阪市中央区淡路町4-3-6: 創元社、2019年2月20日、141–147頁。ISBN 978-4-422-43028-7。
(33)^ 北村雄一︵著︶﹃21世紀こども百科 恐竜館﹄真鍋真︵監修︶、小学館、2007年7月11日、100–101頁。ISBN 978-4092212510。
(34)^ Phil R. Bell; Nicolás E. Campione; W. Scott Persons; Philip J. Currie; Peter L. Larson; Darren H. Tanke; Robert T. Bakker (2017年6月7日). "Tyrannosauroid integument reveals conflicting patterns of gigantism and feather evolution". Biology Letters. 13. doi:10.1098/rsbl.2017.0092。
(35)^ 川上和人﹃鳥類学者 無謀にも恐竜を語る (生物ミステリー)﹄技術評論社、2013年、76頁。ISBN 978-4774155654。
(36)^ abc鹿間時夫﹃古脊椎動物図鑑﹄朝倉書店、1979年、64頁。ISBN 978-4254162226。
(37)^ ab福田芳生﹃図解 恐竜はどんな生物だったか その素顔と生活をさぐる﹄講談社、1987年1月20日、87-88頁。ISBN 9784061326750。
(38)^ Cau, Andrea (13 dicembre 2023). “Theropoda: Tyrannosaurus era verde”. Theropoda. 2024年1月11日閲覧。
(39)^ Currie, P. J. (1998年). "Possible evidence of gregarious behavior in tyrannosaurids". Gaia. 15: 271–277. doi:10.7939/R3348GX03。
(40)^ Joseph E. Peterson; Karsen N. Daus (2019年3月4日). "Feeding traces attributable to juvenile Tyrannosaurus rex offer insight into ontogenetic dietary trends". PeerJ.
(41)^ abMichael Greshko (2020年2月1日). “成長期のティラノサウルス発見 新種論争に決着か”. Nikkei Style. 日本経済新聞社・日経BP. 2020年12月19日閲覧。
(42)^ “ティラノサウルス、これまでの想像より食欲旺盛=研究”. ロイター (2011年10月13日). 2020年12月19日閲覧。
(43)^ ab“Live fast, die young: How dinosaurs lived viciously short lives” (英語). www.sciencefocus.com. 2024年2月23日閲覧。
(44)^ Richard A.Thulborn (1982年1月21日). "Speeds and gaits of dinosaurs". Palaeogeography, Palaeoclimatology, Palaeoecology. 38(3–4): 227–256. doi:10.1016/0031-0182(82)90005-0。
(45)^ James O Farlow; Matt B Smith; John M Robinson (1995年). "Body mass, bone "strength indicator," and cursorial potential of Tyrannosaurus rex". Journal of Vertebrate Paleontology. 15(4): 713–725. doi:10.1080/02724634.1995.10011257。
(46)^ ab﹁ティラノサウルスは俊足? 生体力学で迫る恐竜の走り﹂﹃日本経済新聞﹄2019年7月27日。2020年7月20日閲覧。
(47)^ abc真鍋真、對比地孝亘、ダニエル・L・ブリンクマン、ヒシグシャフ・ツォクバートルイ・ユンナム、小林快次、ツォクトバートル・チンゾリグ、田中康平、今井拓哉、ダーラ・ゼレニスキー、フランソワ・テリエン、グレゴリー・ファンストン、石垣忍、西村智弘、佐藤たまき、小西卓哉、新村龍也、小原正顕、小松俊文、タイラー・R・ライソン、イアン・ミラー、フェルナンド・E・ノバス 著、坂田智佐子 編﹃恐竜博2019 THE DINOSAUR EXPO﹄NHK、NHKプロモーション、朝日新聞社、2019年、146-147頁。
(48)^ Patrick M O'Connor; Leon P A M Claessens (2005年). "Basic avian pulmonary design and flow-through ventilation in non-avian theropod dinosaurs". Nature. doi:10.1038/nature03716。
(49)^ Bill Sellers. “Tyrannosaurus Simulation”. 2005年12月25日時点のオリジナルよりアーカイブ。2020年7月20日閲覧。
(50)^ abSean D.Smith; W. Scott Persons IV; Lida Xingc (2016年). "A tyrannosaur trackway at Glenrock, Lance Formation (Maastrichtian), Wyoming". Cretaceous Research. 61: 1–4.
(51)^ “T. rex was a champion walker, super-efficient at lower speeds”. マギル大学 (2020年5月13日). 2020年7月19日閲覧。
(52)^ abcdBrian Switek (2015年11月11日). “T・レックスのメニュー拝見、ときには共食いも”. ナショナルジオグラフィック協会. 2020年12月19日閲覧。
(53)^ abL.M. Lambe (1917年1月1日). "The Cretaceous Theropodous dinosaur Gorgosaurus". Memoir 100. Geological Survey of Canada. doi:10.4095/101672。
(七)^ “ティランノサウルス・レックス︵動刻︶”. 群馬県立自然史博物館. 2020年12月19日閲覧。
(八)^ 土屋健︵著︶﹃リアルサイズ古生物図鑑 中生代編﹄群馬県立自然史博物館︵監修︶、技術評論社、2019年8月3日、248–251頁。ISBN 978-4-297-10656-0。
(九)^ ヒサクニヒコ﹃新・恐竜論 地球の忘れものを理解する本﹄PHP研究所、2004年2月23日、50頁。ISBN 978-4-569-63334-3。
(十)^ Rebuilding T .Rex - カーネギー自然史博物館のサイトのトピック。
(11)^ abcdef真鍋真、樽創﹁Tyrannosaurus rex 発見百周年﹂﹃化石﹄第72巻、日本古生物学会、2002年9月20日、45–47頁。doi:10.14825/kaseki.72.0_45。2020年12月19日閲覧。
(12)^ Osborn, H.F (1906年). "Tyrannosaurus, Upper Cretaceous carnivorous dinosaur (second communication)". Bulletin of the AMNH. 22: 281.
(13)^ ﹁<中国暴竜>ティラノサウルス類の起源は東アジア、北米移住説覆す―地質科学院﹂﹃Record China﹄2009年10月6日。2020年7月19日閲覧。
(14)^ 動物命名法解説
(15)^ ICZN : Palaeontology & Biostratigraphy
(16)^ “鳥と恐竜は近縁、6800万年前の化石から証拠発見 - 米国”. www.afpbb.com (2007年4月13日). 2023年10月27日閲覧。
(17)^ “利用案内・情報 ≫ ホットニュース ≫ 2009-05-15 :: 国立科学博物館 National Museum of Nature and Science,Tokyo”. www.kahaku.go.jp. 2023年10月27日閲覧。
(18)^ “鳥と恐竜を結び付けたティラノサウルスのタンパク質”. natgeo.nikkeibp.co.jp. 2023年10月27日閲覧。
(19)^ Bath, University of. “New research shows 'juvenile' T. rex fossils are a distinct species of small tyrannosaur” (英語). phys.org. 2024年1月4日閲覧。
(20)^ Longrich, Nicholas R.; Saitta, Evan T. (2024年3月). "Taxonomic Status of Nanotyrannus lancensis (Dinosauria: Tyrannosauroidea)—A Distinct Taxon of Small-Bodied Tyrannosaur". Fossil Studies (英語). 2(1): 1–65. doi:10.3390/fossils2010001. ISSN 2813-6284。
(21)^ abpublished, Sascha Pare (2024年1月3日). “Nanotyrannus vs. T. rex saga continues: Controversial study 'doesn't settle the question at all'” (英語). livescience.com. 2024年1月4日閲覧。
(22)^ Voris, Jared T.; Therrien, Francois; Zelenitzky, Darla K.; Brown, Caleb M. (2020年). "A new tyrannosaurine (Theropoda:Tyrannosauridae) from the Campanian Foremost Formation of Alberta, Canada, provides insight into the evolution and biogeography of tyrannosaurids". Cretaceous Research. 110: 104388. doi:10.1016/j.cretres.2020.104388. S2CID 213838772。
(23)^ abcdefghijデイヴィッド・E・ファストヴスキー [ドイツ語版]、デイヴィッド・B・ウェイシャンペル [英語版]︵著︶﹃恐竜学入門 ─かたち・生態・絶滅─﹄藤原慎一・松本涼子︵訳︶、真鍋真監訳︵第2版版︶、東京都文京区千石3丁目36-7: 東京化学同人、2018年5月25日︵原著2015年1月31日︶、185–190頁。ISBN 978-4-8079-0856-1。NCID BB17901360。OCLC 904989280。全国書誌番号:22535495、ASIN 4807908561。
(24)^ Kent A Stevens (2006年). "Binocular vision in theropod dinosaurs". Journal of Vertebrate Paleontology. 26(2). doi:10.1671/0272-4634(2006)26[321:BVITD]2.0.CO;2。
(25)^ https://www.edu.city.kyoto.jp/science/about/0212tyranno.pdf
(26)^ Lawrence M. Witmer; Ryan C. Ridgely (2009年8月26日). "New Insights Into the Brain, Braincase, and Ear Region of Tyrannosaurs (Dinosauria, Theropoda), with Implications for Sensory Organization and Behavior". The Anatomical Record. 292 (9). doi:10.1002/ar.20983。
(27)^ Martin Kundrát; Xing Xu; Martina Hančová; Andrej Gajdoš; Yu Guo; Defeng Chen (2018年). "Evolutionary disparity in the endoneurocranial configuration between small and gigantic tyrannosauroids". Historical Biology. 32(5): 620–634. doi:10.1080/08912963.2018.1518442。
(28)^ Darla K Zelenitsky; François Therrien; Yoshitsugu Kobayashi (2008年10月28日). "Olfactory acuity in theropods: palaeobiological and evolutionary implications". Proceedings of the Royal Society B. 276 (1657). doi:10.1098/rspb.2008.1075。
(29)^ Thomas D. Carr; David J Varricchio; Jayc C Sedlmayr; Eric M Roberts; Jason R Moore (2017年). "A new tyrannosaur with evidence for anagenesis and crocodile-like facial sensory system". Scientific Reports. 7. doi:10.1038/srep44942。
(30)^ “ティラノサウルスはこんな顔だった、最新報告”. ナショナル・ジオグラフィック協会 (2017年4月4日). 2020年7月20日閲覧。
(31)^ S.V. Saveliev; V. R. Alifanov (2007年). "A new study of the brain of the predatory dinosaur Tarbosaurus bataar (Theropoda, Tyrannosauridae". Paleontological Journal. 41: 281–289. doi:10.1134/S0031030107030070。
(32)^ abcdeダレン・ナイシュ、ポール・バレット︵著︶﹃恐竜の教科書 最新研究で読み解く進化の謎﹄吉田三知世︵訳︶、小林快次・久保田克博・千葉謙太郎・田中康平監訳︵第1版版︶、大阪府大阪市中央区淡路町4-3-6: 創元社、2019年2月20日、141–147頁。ISBN 978-4-422-43028-7。
(33)^ 北村雄一︵著︶﹃21世紀こども百科 恐竜館﹄真鍋真︵監修︶、小学館、2007年7月11日、100–101頁。ISBN 978-4092212510。
(34)^ Phil R. Bell; Nicolás E. Campione; W. Scott Persons; Philip J. Currie; Peter L. Larson; Darren H. Tanke; Robert T. Bakker (2017年6月7日). "Tyrannosauroid integument reveals conflicting patterns of gigantism and feather evolution". Biology Letters. 13. doi:10.1098/rsbl.2017.0092。
(35)^ 川上和人﹃鳥類学者 無謀にも恐竜を語る (生物ミステリー)﹄技術評論社、2013年、76頁。ISBN 978-4774155654。
(36)^ abc鹿間時夫﹃古脊椎動物図鑑﹄朝倉書店、1979年、64頁。ISBN 978-4254162226。
(37)^ ab福田芳生﹃図解 恐竜はどんな生物だったか その素顔と生活をさぐる﹄講談社、1987年1月20日、87-88頁。ISBN 9784061326750。
(38)^ Cau, Andrea (13 dicembre 2023). “Theropoda: Tyrannosaurus era verde”. Theropoda. 2024年1月11日閲覧。
(39)^ Currie, P. J. (1998年). "Possible evidence of gregarious behavior in tyrannosaurids". Gaia. 15: 271–277. doi:10.7939/R3348GX03。
(40)^ Joseph E. Peterson; Karsen N. Daus (2019年3月4日). "Feeding traces attributable to juvenile Tyrannosaurus rex offer insight into ontogenetic dietary trends". PeerJ.
(41)^ abMichael Greshko (2020年2月1日). “成長期のティラノサウルス発見 新種論争に決着か”. Nikkei Style. 日本経済新聞社・日経BP. 2020年12月19日閲覧。
(42)^ “ティラノサウルス、これまでの想像より食欲旺盛=研究”. ロイター (2011年10月13日). 2020年12月19日閲覧。
(43)^ ab“Live fast, die young: How dinosaurs lived viciously short lives” (英語). www.sciencefocus.com. 2024年2月23日閲覧。
(44)^ Richard A.Thulborn (1982年1月21日). "Speeds and gaits of dinosaurs". Palaeogeography, Palaeoclimatology, Palaeoecology. 38(3–4): 227–256. doi:10.1016/0031-0182(82)90005-0。
(45)^ James O Farlow; Matt B Smith; John M Robinson (1995年). "Body mass, bone "strength indicator," and cursorial potential of Tyrannosaurus rex". Journal of Vertebrate Paleontology. 15(4): 713–725. doi:10.1080/02724634.1995.10011257。
(46)^ ab﹁ティラノサウルスは俊足? 生体力学で迫る恐竜の走り﹂﹃日本経済新聞﹄2019年7月27日。2020年7月20日閲覧。
(47)^ abc真鍋真、對比地孝亘、ダニエル・L・ブリンクマン、ヒシグシャフ・ツォクバートルイ・ユンナム、小林快次、ツォクトバートル・チンゾリグ、田中康平、今井拓哉、ダーラ・ゼレニスキー、フランソワ・テリエン、グレゴリー・ファンストン、石垣忍、西村智弘、佐藤たまき、小西卓哉、新村龍也、小原正顕、小松俊文、タイラー・R・ライソン、イアン・ミラー、フェルナンド・E・ノバス 著、坂田智佐子 編﹃恐竜博2019 THE DINOSAUR EXPO﹄NHK、NHKプロモーション、朝日新聞社、2019年、146-147頁。
(48)^ Patrick M O'Connor; Leon P A M Claessens (2005年). "Basic avian pulmonary design and flow-through ventilation in non-avian theropod dinosaurs". Nature. doi:10.1038/nature03716。
(49)^ Bill Sellers. “Tyrannosaurus Simulation”. 2005年12月25日時点のオリジナルよりアーカイブ。2020年7月20日閲覧。
(50)^ abSean D.Smith; W. Scott Persons IV; Lida Xingc (2016年). "A tyrannosaur trackway at Glenrock, Lance Formation (Maastrichtian), Wyoming". Cretaceous Research. 61: 1–4.
(51)^ “T. rex was a champion walker, super-efficient at lower speeds”. マギル大学 (2020年5月13日). 2020年7月19日閲覧。
(52)^ abcdBrian Switek (2015年11月11日). “T・レックスのメニュー拝見、ときには共食いも”. ナショナルジオグラフィック協会. 2020年12月19日閲覧。
(53)^ abL.M. Lambe (1917年1月1日). "The Cretaceous Theropodous dinosaur Gorgosaurus". Memoir 100. Geological Survey of Canada. doi:10.4095/101672。 (54)^ Snyder, Noel F. R. & Helen Snyder (2006). Raptors of North America: Natural History and Conservation. Voyageur Press. p. 40. ISBN 0-7603-2582-0.
(55)^ abcdefgキム・ドユン︵著︶﹃マンガで学ぶ 恐竜の生態﹄渡辺麻土香︵訳︶、マイナビ出版、2020年5月27日、31–58頁。ISBN 978-4-8399-7105-2。
(56)^ abcChris Carbone; Samuel T. Turvey; Jon Bielby (2011年1月26日). "Intra-guild competition and its implications for one of the biggest terrestrial predators, Tyrannosaurus rex". Proc. R. Soc. B. 278 (1718): 2682–2690. doi:10.1098/rspb.2010.2497。
(57)^ Creel, S.; Spong, G.; Creel, N. (2001). "Interspecific competition and the population biology of extinction-prone carnivores". In Gittleman, J. L.; Funk, S. M.; Macdonald, D. W.; Wayne, R. K. (eds.). Carnivore Conservation (1st ed.). Cambridge University Press. pp. 35−60. ISBN 978-0-521-66232-1.
(58)^ Tyrants of the Cenozoic: Evolution of Bone-Crushing Hyenas and Dogs – YouTube
(59)^ Robert A. DePalma II; David A. Burnham; Larry D. Martin; Bruce M. Rothschild; Peter L. Larson (2013年7月30日). "Physical evidence of predatory behavior in Tyrannosaurus rex". PNAS. 30(31). doi:10.1073/pnas.1216534110。
(60)^ Darren Tanke; Bruce Rothschild (2014年1月). Paleopathology in Late Cretaceous Hadrosauridae from Alberta, Canada (Report).
(61)^ Ker Than (2013年3月1日). “T・レックスから逃れたカモノハシ恐竜”. ナショナルジオグラフィック協会. 2016年7月30日時点のオリジナルよりアーカイブ。2020年12月19日閲覧。
(62)^ Kerry Sheridan﹁T・レックスは生きた獲物を捕食、研究﹂ フランス通信社、2013年7月6日。2020年11月27日閲覧。
(63)^ ﹃TATE MUSEUM’S “LEE REX” GETS A NEW HOME(GALLERY)﹄
https://oilcity.news/community/science/2017/05/31/tate-museums-lee-rex-gets-new-home-gallery/
(OIL CITY NEWs:2017)
(64)^ Karen Chin; Tim Tokaryk; Gregory M. Erickson; Lewis C. Calk (1998年). "A king-size theropod coprolite". Nature. 393 (6686): 680–682. doi:10.1038/31461。
(65)^ Holly N. Woodward; Katie Tremaine; Scott A. Williams; Lindsay E. Zanno; John R. Horner; Nathan Myhrvold (2020年1月1日). "Growing up Tyrannosaurus rex: Osteohistology refutes the pygmy "Nanotyrannus" and supports ontogenetic niche partitioning in juvenile Tyrannosaurus". Science Advances. 6(1). doi:10.1126/sciadv.aax6250。
(66)^ Wolff, E. D; Salisbury, S. W; Horner, J. R; Varricchio, D. J (2009年9月30日). "Common avian infection plagued the tyrant dinosaurs". PLOS ONE.
(67)^ “﹁恐竜﹂も新型コロナのような﹁呼吸器感染症﹂にかかっていた︵石田雅彦︶ - エキスパート”. Yahoo!ニュース. 2023年9月1日閲覧。
(68)^ “無敵のT・レックスは寄生虫に負けた?”. natgeo.nikkeibp.co.jp. 2023年9月1日閲覧。
(69)^ “Tレックス最大の敵は寄生虫だった?米豪研究”. www.afpbb.com (2009年10月1日). 2023年9月1日閲覧。
(70)^ Cau, Andrea (29 settembre 2009). “Theropoda: CSI - Hell Creek: Chi uccise Sue?”. Theropoda. 2023年10月11日閲覧。
(71)^ Rothschild, Bruce; O'Connor, Jingmai; Lozado, María Cecilia (2022年12月1日). "Closer examination does not support infection as cause for enigmatic Tyrannosaurus rex mandibular pathologies". Cretaceous Research. 140: 105353. doi:10.1016/j.cretres.2022.105353. ISSN 0195-6671。
(72)^ “All About Stan”. ニューメキシコ自然史科学博物館. 2019年4月4日時点のオリジナルよりアーカイブ。2023年10月31日閲覧。
(73)^ Miller, Jared (2004年5月7日). “Finger bone, details give dino display its edge”. Greatfallstribune.com. 2011年6月29日時点のオリジナルよりアーカイブ。2023年10月31日閲覧。
(74)^ “ギガ恐竜展2017‐地球の絶対王者のなぞ‐ 写真特集‥時事ドットコム”. 時事ドットコム. 2023年11月20日閲覧。
(75)^ ﹃小学館の図鑑NEO 恐竜﹄小学館、2008年6月10日、52−53頁。ISBN 9784092172111。
(76)^ ab北村雄一﹃ティラノサウルス全百科﹄真鍋真︵監修︶、小学館、2005年5月10日、78-79,96-97頁。
(77)^ nandi (2018年1月2日). “Stan the Tyrannosaurus rex”. Paleontology World. 2023年10月31日閲覧。
(78)^ “Unearthing T.Rex”. Black Hills Institute. 2023年10月31日閲覧。
(79)^ Editor, Sanya Nayeem, Games. “Today's Word Search: The accidental discovery of the chicken's ancestor - the T-rex” (英語). Gulf News. 2023年10月31日閲覧。
(80)^ Schweitzer, Mary Higby; Zheng, Wenxia; Zanno, Lindsay; Werning, Sarah; Sugiyama, Toshie (2016年3月15日). "Chemistry supports the identification of gender-specific reproductive tissue in Tyrannosaurus rex". Scientific Reports (英語). 6(1): 23099. doi:10.1038/srep23099. ISSN 2045-2322。
(81)^ “恐竜における骨髄骨の存在を化学的に証明しました | 研究成果 | ニュース”. 新潟大学. 2023年10月31日閲覧。
(82)^ https://jscience.jp/files/docs/119987.pdf
(83)^ “恐竜のオスとメスの違いはわかるの?|恐竜・古生物 Q&A|FPDM: 福井県立恐竜博物館”. FPDM: 福井県立恐竜博物館. 2023年10月31日閲覧。
(84)^ “利用案内・情報 ≫ ホットニュース ≫ 2009-05-15 :: 国立科学博物館 National Museum of Nature and Science,Tokyo”. www.kahaku.go.jp. 2023年11月2日閲覧。
(85)^ "Pregnant T-rex discovery sheds light on evolution of egg-laying". ABC News (オーストラリア英語). 2016年3月16日. 2023年11月2日閲覧。
(86)^ 富田 幸光﹃カラー版 恐竜たちの地球﹄岩波新書、1999年9月29日、137頁。ISBN 978-4004306375。
(87)^ “﹁︻朝日新聞記事から︼ティラノ、脳の遺産 脳囲む骨、日本初公開 ﹂asahi.com : 朝日新聞社 - ﹁恐竜博2011﹂ - ニュース”. www.asahi.com. 2023年10月26日閲覧。
(88)^ ジャン=ギィ・ミシャール﹃恐竜のすべて (﹁知の再発見﹂双書)﹄創元社、1992年、103頁。ISBN 9784422210650。
(89)^ Gignac, P.M; Erickson, G.M (2017年5月17日). "The Biomechanics Behind Extreme Osteophagy in Tyrannosaurus rex". Sci Rep 7. doi:10.1038/s41598-017-02161-w。
(90)^ ab“刷新される恐竜像と私たち~21世紀の恐竜番組~”. NHK. 2020年12月19日閲覧。
(91)^ Paul M.Barrett; Emily J.Rayfield (2006年). "Ecological and evolutionary implications of dinosaur feeding behaviour". Trends in Ecology & Evolution. 21(4): 217–224. doi:10.1016/j.tree.2006.01.002。
(92)^ abBrochu, Christopher A. (2003年1月14日). "Osteology of Tyrannosaurus rex: Insights from a Nearly Complete Skeleton and High-Resolution Computed Tomographic Analysis of the Skull". Memoir. Society of Vertebrate Paleontology. 7. doi:10.2307/3889334. JSTOR 3889334. 2022年10月11日時点のオリジナルよりアーカイブ。2023年11月19日閲覧。
(93)^ Smith, Joshua B. (2005年12月30日). "Heterodonty in Tyrannosaurus rex: implications for the taxonomic and systematic utility of theropod dentitions". Journal of Vertebrate Paleontology (英語). 25(4): 865–887. doi:10.1671/0272-4634(2005)025[0865:HITRIF]2.0.CO;2. ISSN 0272-4634。
(94)^ クリストファー・ロイド﹃137億年の物語‥宇宙が始まってから今日までの全歴史﹄文藝春秋、2012年、68頁。ISBN 978-4163742007。
(95)^ “獲物を襲った巨大な歯 - 日経サイエンス”. www.nikkei-science.com (1999年11月1日). 2024年6月2日閲覧。
(96)^ “Tyrannosaurs softies with offspring, killer instinct with prey | The Asahi Shimbun: Breaking News, Japan News and Analysis” (英語). The Asahi Shimbun. 2023年10月26日閲覧。
(97)^ NEWS, KYODO. “T-Rex may have used lower jaw as tactile sensor: researchers”. Kyodo News+. 2023年10月26日閲覧。
(98)^ ab“ティラノサウルス、甘がみして子ども運んだ?…あご先に高感度の触覚センサー”. 読売新聞オンライン (2021年8月24日). 2023年10月26日閲覧。
(99)^ “T・レックスは舌を突き出せなかった? 研究”. www.afpbb.com (2018年6月21日). 2023年10月31日閲覧。
(100)^ “ティラノ頭骨の謎の穴に新説、頭を冷やす仕組みか”. natgeo.nikkeibp.co.jp. 2023年10月5日閲覧。
(101)^ Holliday, Casey M.; Porter, William Ruger; Vliet, Kent A.; Witmer, Lawrence M. (2020年4月). "The Frontoparietal Fossa and Dorsotemporal Fenestra of Archosaurs and Their Significance for Interpretations of Vascular and Muscular Anatomy in Dinosaurs". The Anatomical Record (英語). 303 (4): 1060–1074. doi:10.1002/ar.24218. ISSN 1932-8486。
(102)^ “人間の脳よりも大きい脳を持つ恐竜はいるの?|恐竜・古生物 Q&A|FPDM: 福井県立恐竜博物館”. FPDM: 福井県立恐竜博物館. 2023年10月26日閲覧。
(103)^ https://www.biorxiv.org/content/10.1101/2022.06.20.496834v1
(104)^ Herculano‐Houzel, Suzana (2023年6月). "Theropod dinosaurs had primate‐like numbers of telencephalic neurons". Journal of Comparative Neurology (英語). 531 (9): 962–974. doi:10.1002/cne.25453. ISSN 0021-9967。
(105)^ https://www.biorxiv.org/content/10.1101/2024.01.10.575006v2
(106)^ マイケル・J・ベントン他︵編︶、2010年﹃生物の進化大図鑑﹄322−323頁。ISBN 978-4-309-25238-4。
(107)^ “霸王龙和近亲用大眼睛换取大咬力—新闻—科学网”. news.sciencenet.cn. 2023年10月26日閲覧。
(108)^ Lautenschlager, Stephan (2022年8月11日). "Functional and ecomorphological evolution of orbit shape in mesozoic archosaurs is driven by body size and diet". Communications Biology (英語). 5(1): 1–11. doi:10.1038/s42003-022-03706-0. ISSN 2399-3642。
(109)^ “古生物学‥ティラノサウルス・レックスとその近縁種は大きな目の代わりに強い咬合力を得た | Communications Biology | Nature Portfolio”. www.natureasia.com. 2023年11月1日閲覧。
(110)^ Cau, Andrea (26 settembre 2021). “Theropoda: L'occhio dei dinosauri”. Theropoda. 2023年11月1日閲覧。
(111)^ Cau, Andrea (26 settembre 2021). “Theropoda: La pupilla dei dinosauri”. Theropoda. 2023年10月11日閲覧。
(112)^ “歯むき出しの肉食恐竜、間違い? ティラノサウルス﹁唇に覆われていた﹂研究成果‥朝日新聞デジタル”. 朝日新聞デジタル (2023年4月19日). 2023年9月1日閲覧。
(113)^ “ティラノサウルスに﹁唇﹂があった可能性、なぜ唇が重要なのか”. natgeo.nikkeibp.co.jp. 2023年9月1日閲覧。
(114)^ “歯は隠れていた?T・レックスに﹁唇﹂あった可能性 米研究”. www.afpbb.com (2023年3月31日). 2023年9月1日閲覧。
(115)^ “歯むき出しのティラノサウルス間違い? 恐竜映画のイメージ覆す研究‥朝日新聞デジタル”. 朝日新聞デジタル (2023年4月13日). 2023年9月1日閲覧。
(116)^ Cullen, Thomas M.; Larson, Derek W.; Witton, Mark P.; Scott, Diane; Maho, Tea; Brink, Kirstin S.; Evans, David C.; Reisz, Robert (2023年3月31日). "Theropod dinosaur facial reconstruction and the importance of soft tissues in paleobiology". Science (英語). 379 (6639): 1348–1352. doi:10.1126/science.abo7877. ISSN 0036-8075。
(117)^ “ティラノサウルスの顔が変わる? | Nature ダイジェスト | Nature Portfolio”. www.natureasia.com. 2023年10月5日閲覧。
(118)^ abG. Masukawa﹃ディノペディア Dinopedia: 恐竜好きのためのイラスト大百科﹄誠文堂新光社、2023年、28-29頁。ISBN 978-4416623510。
(119)^ Carpenter, K. (1997). “Tyrannosauridae”. Encyclopedia of Dinosaurs. San Diego, CA: Academic Press. pp. 766–768. ISBN 978-0-12-226810-6
(120)^ Carpenter, K. (1999). Eggs, Nests, and Baby Dinosaurs: A Look at Dinosaur Reproduction. Bloomington, IN: Indiana University Press. pp. 60–61. ISBN 0-253-33497-7
(121)^ “The anecdotal Tarbosaurus throat skin” (英語). Incertae Sedis (2019年10月6日). 2023年10月16日閲覧。
(122)^ ライリー・ブラック﹃ナショナル ジオグラフィック 世界一美しい恐竜図鑑﹄日経ナショナル ジオグラフィック、2023年1月14日、91頁。ISBN 9784863135550。
(123)^ ﹃新ポケット版学研の図鑑﹁恐竜・大昔の生き物﹂﹄学研プラス、2010年4月22日、145頁。ISBN 978-4-05-203212-7。
(124)^ デビッド・ノーマン、アンジェラ・ミルナー﹃恐竜 (ビジュアル博物館)﹄同朋舎出版、1990年10月1日、14頁。ISBN 978-4810409000。
(125)^ abcde日本古生物学会﹃古生物学辞典﹄朝倉書店、2010年、229頁。ISBN 978-4254162653。
(126)^ “Tyrannosauroidea”. www.theropoddatabase.com. 2023年11月1日閲覧。
(127)^ ﹃小学館の図鑑NEO 大むかしの生物﹄小学館、2007年10月22日、111頁。ISBN 978-4092172128。
(128)^ “No.393 展示紹介 ティラノサウルス|富山市科学博物館 Toyama Science Museum”. www.tsm.toyama.toyama.jp. 2023年9月2日閲覧。
(129)^ “実は凶器? ティラノサウルスの短すぎる腕に新説”. natgeo.nikkeibp.co.jp. 2023年9月1日閲覧。
(130)^ “[https://www.app.pan.pl/article/item/app009212021.html Why tyrannosaurid forelimbs were so short:
An integrative hypothesis - Acta Palaeontologica Polonica]” (英語). www.app.pan.pl. 2023年9月1日閲覧。
(131)^ Bob Bakker (2012年5月9日). “WYREX’S FANCY FOOTWORK AND TENDER HANDS: GET TO KNOW THIS TYRANNOSAUR’S SOFTER SIDE”. BEYOND BONES. ヒューストン自然科学博物館. 2021年5月27日閲覧。
(132)^ 益富 寿之助, 濱田 隆士﹃原色化石図鑑 (保育社の原色図鑑48)︵第15刷︶﹄保育社、1966年、133頁。ISBN 978-4586300488。
(133)^ アラン・チャーリッグ (1987). 恐竜は生きている―新しい恐竜の見方 (自然誌選書). どうぶつ社. p. 130. ISBN 978-4886222305
(134)^ 小畠郁生﹃原色ワイド図鑑 恐竜 ﹇2002﹈改訂新版﹄学研研究社、2002年11月30日、171頁。ISBN 9784051521301。
(135)^ Eric Snively; Haley O’Brien; Donald M. Henderson; Heinrich Mallison; Lara A. Surring; Michael E. Burns; Thomas R. Holtz, Jr; Anthony P. Russell; Lawrence M. Witmer; Philip J. Currie; Scott A. Hartman; John R. Cotton (2019年2月21日). "Lower rotational inertia and larger leg muscles indicate more rapid turns in tyrannosaurids than in other large theropods". PeerJ. doi:10.7717/peerj.6432。
(136)^ David A Krauss; J.M. Robinson (2013年). "The biomechanics of a plausible hunting strategy for Tyrannosaurus rex". Tyrannosaurid Paleobiology: 251–264.
(137)^ 土屋健﹃ティラノサウルスはすごい﹄小林快次︵監修︶、文藝春秋、2015年6月19日。生態の項
(138)^ Andrew A Farke; Ewan D. S. Wolff; Darren H. Tanke (2009年1月28日). "Evidence of combat in Triceratops". PLOS ONE. doi:10.1371/journal.pone.0004252。
(139)^ Tanke, D. H.; Farke, A. A. (2006). “Bone resorption, bone lesions, and extracranial fenestrae in ceratopsid dinosaurs: a preliminary assessment”. In Carpenter, K.. Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs. Bloomington: Indiana University Press. pp. 319–347. ISBN 978-0-253-34817-3
(140)^ Farlow, J. O.; Dodson, P. (1975年). "The behavioral significance of frill and horn morphology in ceratopsian dinosaurs". Evolution. 29(2): 353–361. doi:10.2307/2407222. JSTOR 2407222. PMID 28555861。
(141)^ Martin, A. J. (2006). Introduction to the Study of Dinosaurs (Second ed.). Oxford: Blackwell Publishing. pp. 299–300. ISBN 978-1-4051-3413-2
(142)^ “Tyrannosaurus Rex 'hunted in packs'” (英語). The Telegraph (2011年6月22日). 2023年11月20日閲覧。
(143)^ Cau, Andrea (26 maggio 2009). “Theropoda: Miti e Leggende Post-moderne sui Theropodi Mesozoici - Terza Parte: L'ossessione della Caccia di Gruppo, e le sue alternative (molto più) plausibili”. Theropoda. 2023年11月20日閲覧。
(144)^ Rothschild, Bruce M (2015年). "Unexpected behavior in the Cretaceous: tooth-marked bones attributable to tyrannosaur play". Ethology Ecology & Evolution. 27(3): 325–334. doi:10.1080/03949370.2014.928655。
(145)^ “雷克斯霸王龙 凶残之外也懂幽默?” (中国語). BBC 英伦网 (2015年11月5日). 2023年10月26日閲覧。
(146)^ Bob Yirka (2012年10月29日). “Life, and death, of Triceratops: Fossilized tooth marks shed light on T.Rex's gruesome hunting practices”. Phys.org. 2020年12月19日閲覧。
(147)^ 土屋健﹃化石ドラマチック﹄柴原暁彦︵監修︶、ツク之助︵絵︶、イースト・プレス、2020年5月19日、53-55頁。ISBN 9784781618807。
(148)^ Bruce M Rothschild; Darren Tanke; Ken Carpenter (1997年5月22日). "Tyrannosaurs suffered from gout". Nature. 387. doi:10.1038/387357a0。
(149)^ フジテレビトリビア普及委員会﹃トリビアの泉〜へぇの本〜6﹄講談社、2004年、[要ページ番号]頁。ISBN 978-4063527117。
(150)^ “Tyrannosaurus rex runs again: a theoretical analysis of the hypothesis that full-grown large theropods had a locomotory advantage to hunt in a shallow-water environment”. academic.oup.com. 2023年10月9日閲覧。
(151)^ Currie, P. J. (1998年). "Possible evidence of gregarious behavior in tyrannosaurids". Gaia (15): 271–277. doi:10.7939/R3348GX03. ISSN 0871-5424。
(152)^ “BHI/Fossils & Minerals/Dinosaurs and Birds/STAN T. rex”. bhigr.com. 2023年10月31日閲覧。
(153)^ 講談社﹃小学生の大疑問100パ-ト2 (NHK週刊こどもニュース―図解版スペシャル)﹄講談社、1998年、64−65頁。ISBN 978-4062085656。
(154)^ Peterson J.E., Henderson M.D., Scherer R.P., & Vittore C.P., 2009 - Face Biting on a Juvenile Tyrannosaurid and Behavioral Implications. Palaios 24:780-784.
(155)^ Cau, Andrea (2009年11月30日). “Theropoda: Gerarchia e violenza nell'infanzia di _Tyrannosaurus_”. Theropoda. 2023年10月11日閲覧。
(156)^ ab“﹁ハヤブサの目を持つ男﹂小林快次が﹁ティラノ研究は標本がカギ﹂と主張するワケ︵小林 快次︶”. ブルーバックス | 講談社. 2023年11月2日閲覧。
(157)^ Griebeler, Eva M. (2021年11月). Angielczyk, Kenneth (ed.). "Dinosaurian survivorship schedules revisited: new insights from an age‐structured population model". Palaeontology (英語). 64(6): 839–854. doi:10.1111/pala.12576. ISSN 0031-0239。
(158)^ “﹁素顔は優しい?﹂“最強”ティラノサウルス、最新研究で見えてきた“ちょっと意外な”子育て説︵植田 和貴︶ @gendai_biz”. 現代ビジネス (2021年8月15日). 2023年10月26日閲覧。
(159)^ “ティラノの下顎﹁触覚センサー﹂ 福井県立大研が発表‥中日新聞Web”. 中日新聞Web. 2023年10月26日閲覧。
(160)^ “ティラノサウルスの顎先は高感度の触覚センサーだった可能性を明らかにしました|恐竜博物館の調査研究|FPDM: 福井県立恐竜博物館”. www.dinosaur.pref.fukui.jp. 2023年10月26日閲覧。
(161)^ NHK. “刷新される恐竜像と私たち~21世紀の恐竜番組~|番組|NHKアーカイブス”. 刷新される恐竜像と私たち~21世紀の恐竜番組~|番組|NHKアーカイブス. 2024年1月26日閲覧。
(162)^ トーマス・R・ホルツ、ルイス・V・レイ﹃ホルツ博士の最新恐竜辞典﹄小畠郁生︵監訳︶、朝倉書店、2010年。ISBN 978-4-254-16263-9。カルノサウルスの項。
(163)^ NHK﹁恐竜﹂プロジェクト﹃恐竜VSほ乳類1億5千万年の戦い﹄NHK出版、2006年、[要ページ番号]頁。ISBN 978-4-478-86054-0。
(164)^ NHK﹁ポスト恐竜﹂プロジェクト﹃恐竜絶滅 ほ乳類の戦い﹄NHK出版、2010年、[要ページ番号]頁。ISBN 978-4-478-01386-1。
(165)^ ﹁Hadrosaurs﹂ - GoogleBooks
(166)^ “33億円で落札のティラノ全身化石、今後の研究に懸念も”. ナショナルジオグラフィック協会 (2020年10月6日). 2020年11月27日閲覧。
(167)^ ﹁8000万円で落札のティラノサウルス化石、密輸理由に差し押さえ﹂﹃BB NEWS﹄ フランス通信社、2012年6月29日。2020年7月19日閲覧。
(168)^ ﹁ティラノ骨格化石が8000万円で落札、モンゴル政府は提訴﹂﹃Reuters﹄2012年5月22日。2023年10月5日閲覧。
(169)^ “タルボサウルス解説 - 神流町恐竜センター - 2ページ”. dino-nakasato.org. 2023年10月26日閲覧。
(170)^ Carr, Thomas (2020年10月14日). “Tyrannosauroidea central: Tyrannosaurus rex: 115 Years Old & Royally F***ed”. Tyrannosauroidea central. 2023年11月1日閲覧。
(171)^ Carr, Thomas (2020年11月27日). “Tyrannosauroidea central: WHEN MUSEUMS BUY TYRANNOSAURS”. Tyrannosauroidea central. 2023年11月1日閲覧。
(172)^ “ティラノサウルスは実は3種いた、新たな論文が物議、議論白熱”. natgeo.nikkeibp.co.jp. 2023年9月1日閲覧。
(173)^ “ティラノサウルスは3種いた? 新説発表 王に加え皇帝、女王も‥朝日新聞デジタル”. 朝日新聞デジタル (2022年3月8日). 2023年9月1日閲覧。
(174)^ “33億円のティラノサウルスは新種か 恐竜化石の新説めぐり疑問も‥朝日新聞デジタル”. 朝日新聞デジタル (2022年4月28日). 2023年9月1日閲覧。
(175)^ “[1] Tyrannosaurus remains hint at two possible species distinct from T. rex |
Springer — International Publisher]” (英語). www.springer.com. 2023年9月1日閲覧。
(176)^ Elbein, Asher (2022年3月1日). "They Want to Break T. Rex Into 3 Species. Other Paleontologists Aren't Pleased". The New York Times (アメリカ英語). ISSN 0362-4331. 2023年9月1日閲覧。
(177)^ Davis, Nicola; correspondent, Nicola Davis Science (2022年3月1日). "Tyrannosaurus rex may have been three species, scientists say". The Guardian (イギリス英語). ISSN 0261-3077. 2023年9月1日閲覧。
(178)^ Cau, Andrea (2022年3月1日). “Theropoda: La Trinità di Tyrannosaurus”. Theropoda. 2023年10月11日閲覧。
(179)^ Carr, Thomas D.; Napoli, James G.; Brusatte, Stephen L.; Holtz, Thomas R.; Hone, David W. E.; Williamson, Thomas E.; Zanno, Lindsay E. (2022年9月1日). "Insufficient Evidence for Multiple Species of Tyrannosaurus in the Latest Cretaceous of North America: A Comment on "The Tyrant Lizard King, Queen and Emperor: Multiple Lines of Morphological and Stratigraphic Evidence Support Subtle Evolution and Probable Speciation Within the North American Genus Tyrannosaurus"". Evolutionary Biology (英語). 49(3): 327–341. doi:10.1007/s11692-022-09573-1. ISSN 1934-2845。
(180)^ https://www.amnh.org/explore/news-blogs/research-posts/t-rex-one-species
(181)^ 土屋健﹃も〜っと! 恐竜・古生物ビフォーアフター﹄群馬県立自然史博物館︵監修︶、ツク之助︵絵︶、イースト・プレス、2023年6月20日、29-33頁。ISBN 978-4781622101。
(182)^ updated, Laura Geggel last (2022年7月25日). “'Bold theory' that Tyrannosaurus rex is 3 species gets stomped to pieces” (英語). livescience.com. 2024年1月4日閲覧。
(183)^ Cau, Andrea (2022年3月2日). “Theropoda: Tyrannosaurus rex, Tyrannosaurus rex, Tyrannosaurus rex”. Theropoda. 2023年10月11日閲覧。
(184)^ Dalman, Sebastian G.; Loewen, Mark A.; Pyron, R. Alexander; Jasinski, Steven E.; Malinzak, D. Edward; Lucas, Spencer G.; Fiorillo, Anthony R.; Currie, Philip J.; Longrich, Nicholas R. (2024年1月11日). "A giant tyrannosaur from the Campanian–Maastrichtian of southern North America and the evolution of tyrannosaurid gigantism". Scientific Reports (英語). 13(1): 22124. doi:10.1038/s41598-023-47011-0. ISSN 2045-2322。
(185)^ abcMike (2024年1月11日). “Tyrannosaurus mcraeensis a New Tyrannosaurus Species” (英語). blog.everythingdinosaur.com. 2024年1月11日閲覧。
(186)^ “﹁最強﹂ティラノサウルス・レックスの親戚? 新種化石、米国で発見‥朝日新聞デジタル”. 朝日新聞デジタル (2024年1月12日). 2024年1月26日閲覧。
(187)^ News, A. B. C.. “New dinosaur discovery may be the closest relative to Tyrannosaurus rex, scientists say” (英語). ABC News. 2024年1月12日閲覧。
(188)^ “New species of giant Tyrannosaurus predates T. rex by 6 million years” (英語). New Atlas (2024年1月15日). 2024年1月29日閲覧。
(189)^ “ティラノサウルスだらけのレースが競馬場で開催される アメリカ︵アフロ︶”. Yahoo!ニュース. 2023年10月23日閲覧。
(190)^ ab“さいたま・見沼たんぼでティラノサウルスレース開催へ 浦和エリア初”. 浦和経済新聞. 2023年10月23日閲覧。
(191)^ ab“ティラノサウルス“150人”海岸を全力ダッシュ!たつので﹁おそらく﹂兵庫初、人気着ぐるみレース” (Japanese). 神戸新聞NEXT (2023年10月8日). 2023年10月23日閲覧。
(192)^ “米国発祥のティラノサウルスレース 国内でもじわり、40ほどの大会‥朝日新聞デジタル”. 朝日新聞デジタル (2023年5月17日). 2023年10月24日閲覧。
(193)^ “”恐竜”激走 着ぐるみまとう﹁ティラノサウルスレース﹂﹁丹波竜の里﹂で初開催/兵庫・丹波市︵丹波新聞︶”. Yahoo!ニュース. 2023年10月23日閲覧。
(194)^ 日本放送協会. “鰺ヶ沢町でティラノサウルスレース|NHK 青森県のニュース”. NHK NEWS WEB. 2023年10月23日閲覧。
(195)^ “60匹のティラノサウルス、朱雀門前を全力疾走 奈良・平城宮跡公園‥朝日新聞デジタル”. 朝日新聞デジタル (2023年10月15日). 2023年10月23日閲覧。
(196)^ 日本放送協会. “恐竜“ティラノサウルス” 着ぐるみでレース 秋田 三種町|NHK 秋田県のニュース”. NHK NEWS WEB. 2023年10月23日閲覧。
(197)^ “一度走れば誰もがハマる?ティラノサウルスレース、ブームの予感‥朝日新聞デジタル”. 朝日新聞デジタル (2023年5月17日). 2023年10月24日閲覧。
(198)^ “走るティラノサウルス、浜辺を全力で!海外発祥の人気レース、尼寺のツイッター担当が企画 たつの” (Japanese). 神戸新聞NEXT (2023年5月10日). 2023年10月24日閲覧。
(199)^ “ティラノレース、四国で拡大 観光・イベント連動で誘客”. 日本経済新聞 (2023年9月28日). 2023年10月24日閲覧。
(54)^ Snyder, Noel F. R. & Helen Snyder (2006). Raptors of North America: Natural History and Conservation. Voyageur Press. p. 40. ISBN 0-7603-2582-0.
(55)^ abcdefgキム・ドユン︵著︶﹃マンガで学ぶ 恐竜の生態﹄渡辺麻土香︵訳︶、マイナビ出版、2020年5月27日、31–58頁。ISBN 978-4-8399-7105-2。
(56)^ abcChris Carbone; Samuel T. Turvey; Jon Bielby (2011年1月26日). "Intra-guild competition and its implications for one of the biggest terrestrial predators, Tyrannosaurus rex". Proc. R. Soc. B. 278 (1718): 2682–2690. doi:10.1098/rspb.2010.2497。
(57)^ Creel, S.; Spong, G.; Creel, N. (2001). "Interspecific competition and the population biology of extinction-prone carnivores". In Gittleman, J. L.; Funk, S. M.; Macdonald, D. W.; Wayne, R. K. (eds.). Carnivore Conservation (1st ed.). Cambridge University Press. pp. 35−60. ISBN 978-0-521-66232-1.
(58)^ Tyrants of the Cenozoic: Evolution of Bone-Crushing Hyenas and Dogs – YouTube
(59)^ Robert A. DePalma II; David A. Burnham; Larry D. Martin; Bruce M. Rothschild; Peter L. Larson (2013年7月30日). "Physical evidence of predatory behavior in Tyrannosaurus rex". PNAS. 30(31). doi:10.1073/pnas.1216534110。
(60)^ Darren Tanke; Bruce Rothschild (2014年1月). Paleopathology in Late Cretaceous Hadrosauridae from Alberta, Canada (Report).
(61)^ Ker Than (2013年3月1日). “T・レックスから逃れたカモノハシ恐竜”. ナショナルジオグラフィック協会. 2016年7月30日時点のオリジナルよりアーカイブ。2020年12月19日閲覧。
(62)^ Kerry Sheridan﹁T・レックスは生きた獲物を捕食、研究﹂ フランス通信社、2013年7月6日。2020年11月27日閲覧。
(63)^ ﹃TATE MUSEUM’S “LEE REX” GETS A NEW HOME(GALLERY)﹄
https://oilcity.news/community/science/2017/05/31/tate-museums-lee-rex-gets-new-home-gallery/
(OIL CITY NEWs:2017)
(64)^ Karen Chin; Tim Tokaryk; Gregory M. Erickson; Lewis C. Calk (1998年). "A king-size theropod coprolite". Nature. 393 (6686): 680–682. doi:10.1038/31461。
(65)^ Holly N. Woodward; Katie Tremaine; Scott A. Williams; Lindsay E. Zanno; John R. Horner; Nathan Myhrvold (2020年1月1日). "Growing up Tyrannosaurus rex: Osteohistology refutes the pygmy "Nanotyrannus" and supports ontogenetic niche partitioning in juvenile Tyrannosaurus". Science Advances. 6(1). doi:10.1126/sciadv.aax6250。
(66)^ Wolff, E. D; Salisbury, S. W; Horner, J. R; Varricchio, D. J (2009年9月30日). "Common avian infection plagued the tyrant dinosaurs". PLOS ONE.
(67)^ “﹁恐竜﹂も新型コロナのような﹁呼吸器感染症﹂にかかっていた︵石田雅彦︶ - エキスパート”. Yahoo!ニュース. 2023年9月1日閲覧。
(68)^ “無敵のT・レックスは寄生虫に負けた?”. natgeo.nikkeibp.co.jp. 2023年9月1日閲覧。
(69)^ “Tレックス最大の敵は寄生虫だった?米豪研究”. www.afpbb.com (2009年10月1日). 2023年9月1日閲覧。
(70)^ Cau, Andrea (29 settembre 2009). “Theropoda: CSI - Hell Creek: Chi uccise Sue?”. Theropoda. 2023年10月11日閲覧。
(71)^ Rothschild, Bruce; O'Connor, Jingmai; Lozado, María Cecilia (2022年12月1日). "Closer examination does not support infection as cause for enigmatic Tyrannosaurus rex mandibular pathologies". Cretaceous Research. 140: 105353. doi:10.1016/j.cretres.2022.105353. ISSN 0195-6671。
(72)^ “All About Stan”. ニューメキシコ自然史科学博物館. 2019年4月4日時点のオリジナルよりアーカイブ。2023年10月31日閲覧。
(73)^ Miller, Jared (2004年5月7日). “Finger bone, details give dino display its edge”. Greatfallstribune.com. 2011年6月29日時点のオリジナルよりアーカイブ。2023年10月31日閲覧。
(74)^ “ギガ恐竜展2017‐地球の絶対王者のなぞ‐ 写真特集‥時事ドットコム”. 時事ドットコム. 2023年11月20日閲覧。
(75)^ ﹃小学館の図鑑NEO 恐竜﹄小学館、2008年6月10日、52−53頁。ISBN 9784092172111。
(76)^ ab北村雄一﹃ティラノサウルス全百科﹄真鍋真︵監修︶、小学館、2005年5月10日、78-79,96-97頁。
(77)^ nandi (2018年1月2日). “Stan the Tyrannosaurus rex”. Paleontology World. 2023年10月31日閲覧。
(78)^ “Unearthing T.Rex”. Black Hills Institute. 2023年10月31日閲覧。
(79)^ Editor, Sanya Nayeem, Games. “Today's Word Search: The accidental discovery of the chicken's ancestor - the T-rex” (英語). Gulf News. 2023年10月31日閲覧。
(80)^ Schweitzer, Mary Higby; Zheng, Wenxia; Zanno, Lindsay; Werning, Sarah; Sugiyama, Toshie (2016年3月15日). "Chemistry supports the identification of gender-specific reproductive tissue in Tyrannosaurus rex". Scientific Reports (英語). 6(1): 23099. doi:10.1038/srep23099. ISSN 2045-2322。
(81)^ “恐竜における骨髄骨の存在を化学的に証明しました | 研究成果 | ニュース”. 新潟大学. 2023年10月31日閲覧。
(82)^ https://jscience.jp/files/docs/119987.pdf
(83)^ “恐竜のオスとメスの違いはわかるの?|恐竜・古生物 Q&A|FPDM: 福井県立恐竜博物館”. FPDM: 福井県立恐竜博物館. 2023年10月31日閲覧。
(84)^ “利用案内・情報 ≫ ホットニュース ≫ 2009-05-15 :: 国立科学博物館 National Museum of Nature and Science,Tokyo”. www.kahaku.go.jp. 2023年11月2日閲覧。
(85)^ "Pregnant T-rex discovery sheds light on evolution of egg-laying". ABC News (オーストラリア英語). 2016年3月16日. 2023年11月2日閲覧。
(86)^ 富田 幸光﹃カラー版 恐竜たちの地球﹄岩波新書、1999年9月29日、137頁。ISBN 978-4004306375。
(87)^ “﹁︻朝日新聞記事から︼ティラノ、脳の遺産 脳囲む骨、日本初公開 ﹂asahi.com : 朝日新聞社 - ﹁恐竜博2011﹂ - ニュース”. www.asahi.com. 2023年10月26日閲覧。
(88)^ ジャン=ギィ・ミシャール﹃恐竜のすべて (﹁知の再発見﹂双書)﹄創元社、1992年、103頁。ISBN 9784422210650。
(89)^ Gignac, P.M; Erickson, G.M (2017年5月17日). "The Biomechanics Behind Extreme Osteophagy in Tyrannosaurus rex". Sci Rep 7. doi:10.1038/s41598-017-02161-w。
(90)^ ab“刷新される恐竜像と私たち~21世紀の恐竜番組~”. NHK. 2020年12月19日閲覧。
(91)^ Paul M.Barrett; Emily J.Rayfield (2006年). "Ecological and evolutionary implications of dinosaur feeding behaviour". Trends in Ecology & Evolution. 21(4): 217–224. doi:10.1016/j.tree.2006.01.002。
(92)^ abBrochu, Christopher A. (2003年1月14日). "Osteology of Tyrannosaurus rex: Insights from a Nearly Complete Skeleton and High-Resolution Computed Tomographic Analysis of the Skull". Memoir. Society of Vertebrate Paleontology. 7. doi:10.2307/3889334. JSTOR 3889334. 2022年10月11日時点のオリジナルよりアーカイブ。2023年11月19日閲覧。
(93)^ Smith, Joshua B. (2005年12月30日). "Heterodonty in Tyrannosaurus rex: implications for the taxonomic and systematic utility of theropod dentitions". Journal of Vertebrate Paleontology (英語). 25(4): 865–887. doi:10.1671/0272-4634(2005)025[0865:HITRIF]2.0.CO;2. ISSN 0272-4634。
(94)^ クリストファー・ロイド﹃137億年の物語‥宇宙が始まってから今日までの全歴史﹄文藝春秋、2012年、68頁。ISBN 978-4163742007。
(95)^ “獲物を襲った巨大な歯 - 日経サイエンス”. www.nikkei-science.com (1999年11月1日). 2024年6月2日閲覧。
(96)^ “Tyrannosaurs softies with offspring, killer instinct with prey | The Asahi Shimbun: Breaking News, Japan News and Analysis” (英語). The Asahi Shimbun. 2023年10月26日閲覧。
(97)^ NEWS, KYODO. “T-Rex may have used lower jaw as tactile sensor: researchers”. Kyodo News+. 2023年10月26日閲覧。
(98)^ ab“ティラノサウルス、甘がみして子ども運んだ?…あご先に高感度の触覚センサー”. 読売新聞オンライン (2021年8月24日). 2023年10月26日閲覧。
(99)^ “T・レックスは舌を突き出せなかった? 研究”. www.afpbb.com (2018年6月21日). 2023年10月31日閲覧。
(100)^ “ティラノ頭骨の謎の穴に新説、頭を冷やす仕組みか”. natgeo.nikkeibp.co.jp. 2023年10月5日閲覧。
(101)^ Holliday, Casey M.; Porter, William Ruger; Vliet, Kent A.; Witmer, Lawrence M. (2020年4月). "The Frontoparietal Fossa and Dorsotemporal Fenestra of Archosaurs and Their Significance for Interpretations of Vascular and Muscular Anatomy in Dinosaurs". The Anatomical Record (英語). 303 (4): 1060–1074. doi:10.1002/ar.24218. ISSN 1932-8486。
(102)^ “人間の脳よりも大きい脳を持つ恐竜はいるの?|恐竜・古生物 Q&A|FPDM: 福井県立恐竜博物館”. FPDM: 福井県立恐竜博物館. 2023年10月26日閲覧。
(103)^ https://www.biorxiv.org/content/10.1101/2022.06.20.496834v1
(104)^ Herculano‐Houzel, Suzana (2023年6月). "Theropod dinosaurs had primate‐like numbers of telencephalic neurons". Journal of Comparative Neurology (英語). 531 (9): 962–974. doi:10.1002/cne.25453. ISSN 0021-9967。
(105)^ https://www.biorxiv.org/content/10.1101/2024.01.10.575006v2
(106)^ マイケル・J・ベントン他︵編︶、2010年﹃生物の進化大図鑑﹄322−323頁。ISBN 978-4-309-25238-4。
(107)^ “霸王龙和近亲用大眼睛换取大咬力—新闻—科学网”. news.sciencenet.cn. 2023年10月26日閲覧。
(108)^ Lautenschlager, Stephan (2022年8月11日). "Functional and ecomorphological evolution of orbit shape in mesozoic archosaurs is driven by body size and diet". Communications Biology (英語). 5(1): 1–11. doi:10.1038/s42003-022-03706-0. ISSN 2399-3642。
(109)^ “古生物学‥ティラノサウルス・レックスとその近縁種は大きな目の代わりに強い咬合力を得た | Communications Biology | Nature Portfolio”. www.natureasia.com. 2023年11月1日閲覧。
(110)^ Cau, Andrea (26 settembre 2021). “Theropoda: L'occhio dei dinosauri”. Theropoda. 2023年11月1日閲覧。
(111)^ Cau, Andrea (26 settembre 2021). “Theropoda: La pupilla dei dinosauri”. Theropoda. 2023年10月11日閲覧。
(112)^ “歯むき出しの肉食恐竜、間違い? ティラノサウルス﹁唇に覆われていた﹂研究成果‥朝日新聞デジタル”. 朝日新聞デジタル (2023年4月19日). 2023年9月1日閲覧。
(113)^ “ティラノサウルスに﹁唇﹂があった可能性、なぜ唇が重要なのか”. natgeo.nikkeibp.co.jp. 2023年9月1日閲覧。
(114)^ “歯は隠れていた?T・レックスに﹁唇﹂あった可能性 米研究”. www.afpbb.com (2023年3月31日). 2023年9月1日閲覧。
(115)^ “歯むき出しのティラノサウルス間違い? 恐竜映画のイメージ覆す研究‥朝日新聞デジタル”. 朝日新聞デジタル (2023年4月13日). 2023年9月1日閲覧。
(116)^ Cullen, Thomas M.; Larson, Derek W.; Witton, Mark P.; Scott, Diane; Maho, Tea; Brink, Kirstin S.; Evans, David C.; Reisz, Robert (2023年3月31日). "Theropod dinosaur facial reconstruction and the importance of soft tissues in paleobiology". Science (英語). 379 (6639): 1348–1352. doi:10.1126/science.abo7877. ISSN 0036-8075。
(117)^ “ティラノサウルスの顔が変わる? | Nature ダイジェスト | Nature Portfolio”. www.natureasia.com. 2023年10月5日閲覧。
(118)^ abG. Masukawa﹃ディノペディア Dinopedia: 恐竜好きのためのイラスト大百科﹄誠文堂新光社、2023年、28-29頁。ISBN 978-4416623510。
(119)^ Carpenter, K. (1997). “Tyrannosauridae”. Encyclopedia of Dinosaurs. San Diego, CA: Academic Press. pp. 766–768. ISBN 978-0-12-226810-6
(120)^ Carpenter, K. (1999). Eggs, Nests, and Baby Dinosaurs: A Look at Dinosaur Reproduction. Bloomington, IN: Indiana University Press. pp. 60–61. ISBN 0-253-33497-7
(121)^ “The anecdotal Tarbosaurus throat skin” (英語). Incertae Sedis (2019年10月6日). 2023年10月16日閲覧。
(122)^ ライリー・ブラック﹃ナショナル ジオグラフィック 世界一美しい恐竜図鑑﹄日経ナショナル ジオグラフィック、2023年1月14日、91頁。ISBN 9784863135550。
(123)^ ﹃新ポケット版学研の図鑑﹁恐竜・大昔の生き物﹂﹄学研プラス、2010年4月22日、145頁。ISBN 978-4-05-203212-7。
(124)^ デビッド・ノーマン、アンジェラ・ミルナー﹃恐竜 (ビジュアル博物館)﹄同朋舎出版、1990年10月1日、14頁。ISBN 978-4810409000。
(125)^ abcde日本古生物学会﹃古生物学辞典﹄朝倉書店、2010年、229頁。ISBN 978-4254162653。
(126)^ “Tyrannosauroidea”. www.theropoddatabase.com. 2023年11月1日閲覧。
(127)^ ﹃小学館の図鑑NEO 大むかしの生物﹄小学館、2007年10月22日、111頁。ISBN 978-4092172128。
(128)^ “No.393 展示紹介 ティラノサウルス|富山市科学博物館 Toyama Science Museum”. www.tsm.toyama.toyama.jp. 2023年9月2日閲覧。
(129)^ “実は凶器? ティラノサウルスの短すぎる腕に新説”. natgeo.nikkeibp.co.jp. 2023年9月1日閲覧。
(130)^ “[https://www.app.pan.pl/article/item/app009212021.html Why tyrannosaurid forelimbs were so short:
An integrative hypothesis - Acta Palaeontologica Polonica]” (英語). www.app.pan.pl. 2023年9月1日閲覧。
(131)^ Bob Bakker (2012年5月9日). “WYREX’S FANCY FOOTWORK AND TENDER HANDS: GET TO KNOW THIS TYRANNOSAUR’S SOFTER SIDE”. BEYOND BONES. ヒューストン自然科学博物館. 2021年5月27日閲覧。
(132)^ 益富 寿之助, 濱田 隆士﹃原色化石図鑑 (保育社の原色図鑑48)︵第15刷︶﹄保育社、1966年、133頁。ISBN 978-4586300488。
(133)^ アラン・チャーリッグ (1987). 恐竜は生きている―新しい恐竜の見方 (自然誌選書). どうぶつ社. p. 130. ISBN 978-4886222305
(134)^ 小畠郁生﹃原色ワイド図鑑 恐竜 ﹇2002﹈改訂新版﹄学研研究社、2002年11月30日、171頁。ISBN 9784051521301。
(135)^ Eric Snively; Haley O’Brien; Donald M. Henderson; Heinrich Mallison; Lara A. Surring; Michael E. Burns; Thomas R. Holtz, Jr; Anthony P. Russell; Lawrence M. Witmer; Philip J. Currie; Scott A. Hartman; John R. Cotton (2019年2月21日). "Lower rotational inertia and larger leg muscles indicate more rapid turns in tyrannosaurids than in other large theropods". PeerJ. doi:10.7717/peerj.6432。
(136)^ David A Krauss; J.M. Robinson (2013年). "The biomechanics of a plausible hunting strategy for Tyrannosaurus rex". Tyrannosaurid Paleobiology: 251–264.
(137)^ 土屋健﹃ティラノサウルスはすごい﹄小林快次︵監修︶、文藝春秋、2015年6月19日。生態の項
(138)^ Andrew A Farke; Ewan D. S. Wolff; Darren H. Tanke (2009年1月28日). "Evidence of combat in Triceratops". PLOS ONE. doi:10.1371/journal.pone.0004252。
(139)^ Tanke, D. H.; Farke, A. A. (2006). “Bone resorption, bone lesions, and extracranial fenestrae in ceratopsid dinosaurs: a preliminary assessment”. In Carpenter, K.. Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs. Bloomington: Indiana University Press. pp. 319–347. ISBN 978-0-253-34817-3
(140)^ Farlow, J. O.; Dodson, P. (1975年). "The behavioral significance of frill and horn morphology in ceratopsian dinosaurs". Evolution. 29(2): 353–361. doi:10.2307/2407222. JSTOR 2407222. PMID 28555861。
(141)^ Martin, A. J. (2006). Introduction to the Study of Dinosaurs (Second ed.). Oxford: Blackwell Publishing. pp. 299–300. ISBN 978-1-4051-3413-2
(142)^ “Tyrannosaurus Rex 'hunted in packs'” (英語). The Telegraph (2011年6月22日). 2023年11月20日閲覧。
(143)^ Cau, Andrea (26 maggio 2009). “Theropoda: Miti e Leggende Post-moderne sui Theropodi Mesozoici - Terza Parte: L'ossessione della Caccia di Gruppo, e le sue alternative (molto più) plausibili”. Theropoda. 2023年11月20日閲覧。
(144)^ Rothschild, Bruce M (2015年). "Unexpected behavior in the Cretaceous: tooth-marked bones attributable to tyrannosaur play". Ethology Ecology & Evolution. 27(3): 325–334. doi:10.1080/03949370.2014.928655。
(145)^ “雷克斯霸王龙 凶残之外也懂幽默?” (中国語). BBC 英伦网 (2015年11月5日). 2023年10月26日閲覧。
(146)^ Bob Yirka (2012年10月29日). “Life, and death, of Triceratops: Fossilized tooth marks shed light on T.Rex's gruesome hunting practices”. Phys.org. 2020年12月19日閲覧。
(147)^ 土屋健﹃化石ドラマチック﹄柴原暁彦︵監修︶、ツク之助︵絵︶、イースト・プレス、2020年5月19日、53-55頁。ISBN 9784781618807。
(148)^ Bruce M Rothschild; Darren Tanke; Ken Carpenter (1997年5月22日). "Tyrannosaurs suffered from gout". Nature. 387. doi:10.1038/387357a0。
(149)^ フジテレビトリビア普及委員会﹃トリビアの泉〜へぇの本〜6﹄講談社、2004年、[要ページ番号]頁。ISBN 978-4063527117。
(150)^ “Tyrannosaurus rex runs again: a theoretical analysis of the hypothesis that full-grown large theropods had a locomotory advantage to hunt in a shallow-water environment”. academic.oup.com. 2023年10月9日閲覧。
(151)^ Currie, P. J. (1998年). "Possible evidence of gregarious behavior in tyrannosaurids". Gaia (15): 271–277. doi:10.7939/R3348GX03. ISSN 0871-5424。
(152)^ “BHI/Fossils & Minerals/Dinosaurs and Birds/STAN T. rex”. bhigr.com. 2023年10月31日閲覧。
(153)^ 講談社﹃小学生の大疑問100パ-ト2 (NHK週刊こどもニュース―図解版スペシャル)﹄講談社、1998年、64−65頁。ISBN 978-4062085656。
(154)^ Peterson J.E., Henderson M.D., Scherer R.P., & Vittore C.P., 2009 - Face Biting on a Juvenile Tyrannosaurid and Behavioral Implications. Palaios 24:780-784.
(155)^ Cau, Andrea (2009年11月30日). “Theropoda: Gerarchia e violenza nell'infanzia di _Tyrannosaurus_”. Theropoda. 2023年10月11日閲覧。
(156)^ ab“﹁ハヤブサの目を持つ男﹂小林快次が﹁ティラノ研究は標本がカギ﹂と主張するワケ︵小林 快次︶”. ブルーバックス | 講談社. 2023年11月2日閲覧。
(157)^ Griebeler, Eva M. (2021年11月). Angielczyk, Kenneth (ed.). "Dinosaurian survivorship schedules revisited: new insights from an age‐structured population model". Palaeontology (英語). 64(6): 839–854. doi:10.1111/pala.12576. ISSN 0031-0239。
(158)^ “﹁素顔は優しい?﹂“最強”ティラノサウルス、最新研究で見えてきた“ちょっと意外な”子育て説︵植田 和貴︶ @gendai_biz”. 現代ビジネス (2021年8月15日). 2023年10月26日閲覧。
(159)^ “ティラノの下顎﹁触覚センサー﹂ 福井県立大研が発表‥中日新聞Web”. 中日新聞Web. 2023年10月26日閲覧。
(160)^ “ティラノサウルスの顎先は高感度の触覚センサーだった可能性を明らかにしました|恐竜博物館の調査研究|FPDM: 福井県立恐竜博物館”. www.dinosaur.pref.fukui.jp. 2023年10月26日閲覧。
(161)^ NHK. “刷新される恐竜像と私たち~21世紀の恐竜番組~|番組|NHKアーカイブス”. 刷新される恐竜像と私たち~21世紀の恐竜番組~|番組|NHKアーカイブス. 2024年1月26日閲覧。
(162)^ トーマス・R・ホルツ、ルイス・V・レイ﹃ホルツ博士の最新恐竜辞典﹄小畠郁生︵監訳︶、朝倉書店、2010年。ISBN 978-4-254-16263-9。カルノサウルスの項。
(163)^ NHK﹁恐竜﹂プロジェクト﹃恐竜VSほ乳類1億5千万年の戦い﹄NHK出版、2006年、[要ページ番号]頁。ISBN 978-4-478-86054-0。
(164)^ NHK﹁ポスト恐竜﹂プロジェクト﹃恐竜絶滅 ほ乳類の戦い﹄NHK出版、2010年、[要ページ番号]頁。ISBN 978-4-478-01386-1。
(165)^ ﹁Hadrosaurs﹂ - GoogleBooks
(166)^ “33億円で落札のティラノ全身化石、今後の研究に懸念も”. ナショナルジオグラフィック協会 (2020年10月6日). 2020年11月27日閲覧。
(167)^ ﹁8000万円で落札のティラノサウルス化石、密輸理由に差し押さえ﹂﹃BB NEWS﹄ フランス通信社、2012年6月29日。2020年7月19日閲覧。
(168)^ ﹁ティラノ骨格化石が8000万円で落札、モンゴル政府は提訴﹂﹃Reuters﹄2012年5月22日。2023年10月5日閲覧。
(169)^ “タルボサウルス解説 - 神流町恐竜センター - 2ページ”. dino-nakasato.org. 2023年10月26日閲覧。
(170)^ Carr, Thomas (2020年10月14日). “Tyrannosauroidea central: Tyrannosaurus rex: 115 Years Old & Royally F***ed”. Tyrannosauroidea central. 2023年11月1日閲覧。
(171)^ Carr, Thomas (2020年11月27日). “Tyrannosauroidea central: WHEN MUSEUMS BUY TYRANNOSAURS”. Tyrannosauroidea central. 2023年11月1日閲覧。
(172)^ “ティラノサウルスは実は3種いた、新たな論文が物議、議論白熱”. natgeo.nikkeibp.co.jp. 2023年9月1日閲覧。
(173)^ “ティラノサウルスは3種いた? 新説発表 王に加え皇帝、女王も‥朝日新聞デジタル”. 朝日新聞デジタル (2022年3月8日). 2023年9月1日閲覧。
(174)^ “33億円のティラノサウルスは新種か 恐竜化石の新説めぐり疑問も‥朝日新聞デジタル”. 朝日新聞デジタル (2022年4月28日). 2023年9月1日閲覧。
(175)^ “[1] Tyrannosaurus remains hint at two possible species distinct from T. rex |
Springer — International Publisher]” (英語). www.springer.com. 2023年9月1日閲覧。
(176)^ Elbein, Asher (2022年3月1日). "They Want to Break T. Rex Into 3 Species. Other Paleontologists Aren't Pleased". The New York Times (アメリカ英語). ISSN 0362-4331. 2023年9月1日閲覧。
(177)^ Davis, Nicola; correspondent, Nicola Davis Science (2022年3月1日). "Tyrannosaurus rex may have been three species, scientists say". The Guardian (イギリス英語). ISSN 0261-3077. 2023年9月1日閲覧。
(178)^ Cau, Andrea (2022年3月1日). “Theropoda: La Trinità di Tyrannosaurus”. Theropoda. 2023年10月11日閲覧。
(179)^ Carr, Thomas D.; Napoli, James G.; Brusatte, Stephen L.; Holtz, Thomas R.; Hone, David W. E.; Williamson, Thomas E.; Zanno, Lindsay E. (2022年9月1日). "Insufficient Evidence for Multiple Species of Tyrannosaurus in the Latest Cretaceous of North America: A Comment on "The Tyrant Lizard King, Queen and Emperor: Multiple Lines of Morphological and Stratigraphic Evidence Support Subtle Evolution and Probable Speciation Within the North American Genus Tyrannosaurus"". Evolutionary Biology (英語). 49(3): 327–341. doi:10.1007/s11692-022-09573-1. ISSN 1934-2845。
(180)^ https://www.amnh.org/explore/news-blogs/research-posts/t-rex-one-species
(181)^ 土屋健﹃も〜っと! 恐竜・古生物ビフォーアフター﹄群馬県立自然史博物館︵監修︶、ツク之助︵絵︶、イースト・プレス、2023年6月20日、29-33頁。ISBN 978-4781622101。
(182)^ updated, Laura Geggel last (2022年7月25日). “'Bold theory' that Tyrannosaurus rex is 3 species gets stomped to pieces” (英語). livescience.com. 2024年1月4日閲覧。
(183)^ Cau, Andrea (2022年3月2日). “Theropoda: Tyrannosaurus rex, Tyrannosaurus rex, Tyrannosaurus rex”. Theropoda. 2023年10月11日閲覧。
(184)^ Dalman, Sebastian G.; Loewen, Mark A.; Pyron, R. Alexander; Jasinski, Steven E.; Malinzak, D. Edward; Lucas, Spencer G.; Fiorillo, Anthony R.; Currie, Philip J.; Longrich, Nicholas R. (2024年1月11日). "A giant tyrannosaur from the Campanian–Maastrichtian of southern North America and the evolution of tyrannosaurid gigantism". Scientific Reports (英語). 13(1): 22124. doi:10.1038/s41598-023-47011-0. ISSN 2045-2322。
(185)^ abcMike (2024年1月11日). “Tyrannosaurus mcraeensis a New Tyrannosaurus Species” (英語). blog.everythingdinosaur.com. 2024年1月11日閲覧。
(186)^ “﹁最強﹂ティラノサウルス・レックスの親戚? 新種化石、米国で発見‥朝日新聞デジタル”. 朝日新聞デジタル (2024年1月12日). 2024年1月26日閲覧。
(187)^ News, A. B. C.. “New dinosaur discovery may be the closest relative to Tyrannosaurus rex, scientists say” (英語). ABC News. 2024年1月12日閲覧。
(188)^ “New species of giant Tyrannosaurus predates T. rex by 6 million years” (英語). New Atlas (2024年1月15日). 2024年1月29日閲覧。
(189)^ “ティラノサウルスだらけのレースが競馬場で開催される アメリカ︵アフロ︶”. Yahoo!ニュース. 2023年10月23日閲覧。
(190)^ ab“さいたま・見沼たんぼでティラノサウルスレース開催へ 浦和エリア初”. 浦和経済新聞. 2023年10月23日閲覧。
(191)^ ab“ティラノサウルス“150人”海岸を全力ダッシュ!たつので﹁おそらく﹂兵庫初、人気着ぐるみレース” (Japanese). 神戸新聞NEXT (2023年10月8日). 2023年10月23日閲覧。
(192)^ “米国発祥のティラノサウルスレース 国内でもじわり、40ほどの大会‥朝日新聞デジタル”. 朝日新聞デジタル (2023年5月17日). 2023年10月24日閲覧。
(193)^ “”恐竜”激走 着ぐるみまとう﹁ティラノサウルスレース﹂﹁丹波竜の里﹂で初開催/兵庫・丹波市︵丹波新聞︶”. Yahoo!ニュース. 2023年10月23日閲覧。
(194)^ 日本放送協会. “鰺ヶ沢町でティラノサウルスレース|NHK 青森県のニュース”. NHK NEWS WEB. 2023年10月23日閲覧。
(195)^ “60匹のティラノサウルス、朱雀門前を全力疾走 奈良・平城宮跡公園‥朝日新聞デジタル”. 朝日新聞デジタル (2023年10月15日). 2023年10月23日閲覧。
(196)^ 日本放送協会. “恐竜“ティラノサウルス” 着ぐるみでレース 秋田 三種町|NHK 秋田県のニュース”. NHK NEWS WEB. 2023年10月23日閲覧。
(197)^ “一度走れば誰もがハマる?ティラノサウルスレース、ブームの予感‥朝日新聞デジタル”. 朝日新聞デジタル (2023年5月17日). 2023年10月24日閲覧。
(198)^ “走るティラノサウルス、浜辺を全力で!海外発祥の人気レース、尼寺のツイッター担当が企画 たつの” (Japanese). 神戸新聞NEXT (2023年5月10日). 2023年10月24日閲覧。
(199)^ “ティラノレース、四国で拡大 観光・イベント連動で誘客”. 日本経済新聞 (2023年9月28日). 2023年10月24日閲覧。
 (七)^ “ティランノサウルス・レックス︵動刻︶”. 群馬県立自然史博物館. 2020年12月19日閲覧。
(八)^ 土屋健︵著︶﹃リアルサイズ古生物図鑑 中生代編﹄群馬県立自然史博物館︵監修︶、技術評論社、2019年8月3日、248–251頁。ISBN 978-4-297-10656-0。
(九)^ ヒサクニヒコ﹃新・恐竜論 地球の忘れものを理解する本﹄PHP研究所、2004年2月23日、50頁。ISBN 978-4-569-63334-3。
(十)^ Rebuilding T .Rex - カーネギー自然史博物館のサイトのトピック。
(11)^ abcdef真鍋真、樽創﹁Tyrannosaurus rex 発見百周年﹂﹃化石﹄第72巻、日本古生物学会、2002年9月20日、45–47頁。doi:10.14825/kaseki.72.0_45。2020年12月19日閲覧。
(12)^ Osborn, H.F (1906年). "Tyrannosaurus, Upper Cretaceous carnivorous dinosaur (second communication)". Bulletin of the AMNH. 22: 281.
(13)^ ﹁<中国暴竜>ティラノサウルス類の起源は東アジア、北米移住説覆す―地質科学院﹂﹃Record China﹄2009年10月6日。2020年7月19日閲覧。
(14)^ 動物命名法解説
(15)^ ICZN : Palaeontology & Biostratigraphy
(16)^ “鳥と恐竜は近縁、6800万年前の化石から証拠発見 - 米国”. www.afpbb.com (2007年4月13日). 2023年10月27日閲覧。
(17)^ “利用案内・情報 ≫ ホットニュース ≫ 2009-05-15 :: 国立科学博物館 National Museum of Nature and Science,Tokyo”. www.kahaku.go.jp. 2023年10月27日閲覧。
(18)^ “鳥と恐竜を結び付けたティラノサウルスのタンパク質”. natgeo.nikkeibp.co.jp. 2023年10月27日閲覧。
(19)^ Bath, University of. “New research shows 'juvenile' T. rex fossils are a distinct species of small tyrannosaur” (英語). phys.org. 2024年1月4日閲覧。
(20)^ Longrich, Nicholas R.; Saitta, Evan T. (2024年3月). "Taxonomic Status of Nanotyrannus lancensis (Dinosauria: Tyrannosauroidea)—A Distinct Taxon of Small-Bodied Tyrannosaur". Fossil Studies (英語). 2(1): 1–65. doi:10.3390/fossils2010001. ISSN 2813-6284。
(21)^ abpublished, Sascha Pare (2024年1月3日). “Nanotyrannus vs. T. rex saga continues: Controversial study 'doesn't settle the question at all'” (英語). livescience.com. 2024年1月4日閲覧。
(22)^ Voris, Jared T.; Therrien, Francois; Zelenitzky, Darla K.; Brown, Caleb M. (2020年). "A new tyrannosaurine (Theropoda:Tyrannosauridae) from the Campanian Foremost Formation of Alberta, Canada, provides insight into the evolution and biogeography of tyrannosaurids". Cretaceous Research. 110: 104388. doi:10.1016/j.cretres.2020.104388. S2CID 213838772。
(23)^ abcdefghijデイヴィッド・E・ファストヴスキー [ドイツ語版]、デイヴィッド・B・ウェイシャンペル [英語版]︵著︶﹃恐竜学入門 ─かたち・生態・絶滅─﹄藤原慎一・松本涼子︵訳︶、真鍋真監訳︵第2版版︶、東京都文京区千石3丁目36-7: 東京化学同人、2018年5月25日︵原著2015年1月31日︶、185–190頁。ISBN 978-4-8079-0856-1。NCID BB17901360。OCLC 904989280。全国書誌番号:22535495、ASIN 4807908561。
(24)^ Kent A Stevens (2006年). "Binocular vision in theropod dinosaurs". Journal of Vertebrate Paleontology. 26(2). doi:10.1671/0272-4634(2006)26[321:BVITD]2.0.CO;2。
(25)^ https://www.edu.city.kyoto.jp/science/about/0212tyranno.pdf
(26)^ Lawrence M. Witmer; Ryan C. Ridgely (2009年8月26日). "New Insights Into the Brain, Braincase, and Ear Region of Tyrannosaurs (Dinosauria, Theropoda), with Implications for Sensory Organization and Behavior". The Anatomical Record. 292 (9). doi:10.1002/ar.20983。
(27)^ Martin Kundrát; Xing Xu; Martina Hančová; Andrej Gajdoš; Yu Guo; Defeng Chen (2018年). "Evolutionary disparity in the endoneurocranial configuration between small and gigantic tyrannosauroids". Historical Biology. 32(5): 620–634. doi:10.1080/08912963.2018.1518442。
(28)^ Darla K Zelenitsky; François Therrien; Yoshitsugu Kobayashi (2008年10月28日). "Olfactory acuity in theropods: palaeobiological and evolutionary implications". Proceedings of the Royal Society B. 276 (1657). doi:10.1098/rspb.2008.1075。
(29)^ Thomas D. Carr; David J Varricchio; Jayc C Sedlmayr; Eric M Roberts; Jason R Moore (2017年). "A new tyrannosaur with evidence for anagenesis and crocodile-like facial sensory system". Scientific Reports. 7. doi:10.1038/srep44942。
(30)^ “ティラノサウルスはこんな顔だった、最新報告”. ナショナル・ジオグラフィック協会 (2017年4月4日). 2020年7月20日閲覧。
(31)^ S.V. Saveliev; V. R. Alifanov (2007年). "A new study of the brain of the predatory dinosaur Tarbosaurus bataar (Theropoda, Tyrannosauridae". Paleontological Journal. 41: 281–289. doi:10.1134/S0031030107030070。
(32)^ abcdeダレン・ナイシュ、ポール・バレット︵著︶﹃恐竜の教科書 最新研究で読み解く進化の謎﹄吉田三知世︵訳︶、小林快次・久保田克博・千葉謙太郎・田中康平監訳︵第1版版︶、大阪府大阪市中央区淡路町4-3-6: 創元社、2019年2月20日、141–147頁。ISBN 978-4-422-43028-7。
(33)^ 北村雄一︵著︶﹃21世紀こども百科 恐竜館﹄真鍋真︵監修︶、小学館、2007年7月11日、100–101頁。ISBN 978-4092212510。
(34)^ Phil R. Bell; Nicolás E. Campione; W. Scott Persons; Philip J. Currie; Peter L. Larson; Darren H. Tanke; Robert T. Bakker (2017年6月7日). "Tyrannosauroid integument reveals conflicting patterns of gigantism and feather evolution". Biology Letters. 13. doi:10.1098/rsbl.2017.0092。
(35)^ 川上和人﹃鳥類学者 無謀にも恐竜を語る (生物ミステリー)﹄技術評論社、2013年、76頁。ISBN 978-4774155654。
(36)^ abc鹿間時夫﹃古脊椎動物図鑑﹄朝倉書店、1979年、64頁。ISBN 978-4254162226。
(37)^ ab福田芳生﹃図解 恐竜はどんな生物だったか その素顔と生活をさぐる﹄講談社、1987年1月20日、87-88頁。ISBN 9784061326750。
(38)^ Cau, Andrea (13 dicembre 2023). “Theropoda: Tyrannosaurus era verde”. Theropoda. 2024年1月11日閲覧。
(39)^ Currie, P. J. (1998年). "Possible evidence of gregarious behavior in tyrannosaurids". Gaia. 15: 271–277. doi:10.7939/R3348GX03。
(40)^ Joseph E. Peterson; Karsen N. Daus (2019年3月4日). "Feeding traces attributable to juvenile Tyrannosaurus rex offer insight into ontogenetic dietary trends". PeerJ.
(41)^ abMichael Greshko (2020年2月1日). “成長期のティラノサウルス発見 新種論争に決着か”. Nikkei Style. 日本経済新聞社・日経BP. 2020年12月19日閲覧。
(42)^ “ティラノサウルス、これまでの想像より食欲旺盛=研究”. ロイター (2011年10月13日). 2020年12月19日閲覧。
(43)^ ab“Live fast, die young: How dinosaurs lived viciously short lives” (英語). www.sciencefocus.com. 2024年2月23日閲覧。
(44)^ Richard A.Thulborn (1982年1月21日). "Speeds and gaits of dinosaurs". Palaeogeography, Palaeoclimatology, Palaeoecology. 38(3–4): 227–256. doi:10.1016/0031-0182(82)90005-0。
(45)^ James O Farlow; Matt B Smith; John M Robinson (1995年). "Body mass, bone "strength indicator," and cursorial potential of Tyrannosaurus rex". Journal of Vertebrate Paleontology. 15(4): 713–725. doi:10.1080/02724634.1995.10011257。
(46)^ ab﹁ティラノサウルスは俊足? 生体力学で迫る恐竜の走り﹂﹃日本経済新聞﹄2019年7月27日。2020年7月20日閲覧。
(47)^ abc真鍋真、對比地孝亘、ダニエル・L・ブリンクマン、ヒシグシャフ・ツォクバートルイ・ユンナム、小林快次、ツォクトバートル・チンゾリグ、田中康平、今井拓哉、ダーラ・ゼレニスキー、フランソワ・テリエン、グレゴリー・ファンストン、石垣忍、西村智弘、佐藤たまき、小西卓哉、新村龍也、小原正顕、小松俊文、タイラー・R・ライソン、イアン・ミラー、フェルナンド・E・ノバス 著、坂田智佐子 編﹃恐竜博2019 THE DINOSAUR EXPO﹄NHK、NHKプロモーション、朝日新聞社、2019年、146-147頁。
(48)^ Patrick M O'Connor; Leon P A M Claessens (2005年). "Basic avian pulmonary design and flow-through ventilation in non-avian theropod dinosaurs". Nature. doi:10.1038/nature03716。
(49)^ Bill Sellers. “Tyrannosaurus Simulation”. 2005年12月25日時点のオリジナルよりアーカイブ。2020年7月20日閲覧。
(50)^ abSean D.Smith; W. Scott Persons IV; Lida Xingc (2016年). "A tyrannosaur trackway at Glenrock, Lance Formation (Maastrichtian), Wyoming". Cretaceous Research. 61: 1–4.
(51)^ “T. rex was a champion walker, super-efficient at lower speeds”. マギル大学 (2020年5月13日). 2020年7月19日閲覧。
(52)^ abcdBrian Switek (2015年11月11日). “T・レックスのメニュー拝見、ときには共食いも”. ナショナルジオグラフィック協会. 2020年12月19日閲覧。
(53)^ abL.M. Lambe (1917年1月1日). "The Cretaceous Theropodous dinosaur Gorgosaurus". Memoir 100. Geological Survey of Canada. doi:10.4095/101672。
(136)^ David A Krauss; J.M. Robinson (2013年). "The biomechanics of a plausible hunting strategy for Tyrannosaurus rex". Tyrannosaurid Paleobiology: 251–264.
(137)^ 土屋健﹃ティラノサウルスはすごい﹄小林快次︵監修︶、文藝春秋、2015年6月19日。生態の項
(138)^ Andrew A Farke; Ewan D. S. Wolff; Darren H. Tanke (2009年1月28日). "Evidence of combat in Triceratops". PLOS ONE. doi:10.1371/journal.pone.0004252。
(七)^ “ティランノサウルス・レックス︵動刻︶”. 群馬県立自然史博物館. 2020年12月19日閲覧。
(八)^ 土屋健︵著︶﹃リアルサイズ古生物図鑑 中生代編﹄群馬県立自然史博物館︵監修︶、技術評論社、2019年8月3日、248–251頁。ISBN 978-4-297-10656-0。
(九)^ ヒサクニヒコ﹃新・恐竜論 地球の忘れものを理解する本﹄PHP研究所、2004年2月23日、50頁。ISBN 978-4-569-63334-3。
(十)^ Rebuilding T .Rex - カーネギー自然史博物館のサイトのトピック。
(11)^ abcdef真鍋真、樽創﹁Tyrannosaurus rex 発見百周年﹂﹃化石﹄第72巻、日本古生物学会、2002年9月20日、45–47頁。doi:10.14825/kaseki.72.0_45。2020年12月19日閲覧。
(12)^ Osborn, H.F (1906年). "Tyrannosaurus, Upper Cretaceous carnivorous dinosaur (second communication)". Bulletin of the AMNH. 22: 281.
(13)^ ﹁<中国暴竜>ティラノサウルス類の起源は東アジア、北米移住説覆す―地質科学院﹂﹃Record China﹄2009年10月6日。2020年7月19日閲覧。
(14)^ 動物命名法解説
(15)^ ICZN : Palaeontology & Biostratigraphy
(16)^ “鳥と恐竜は近縁、6800万年前の化石から証拠発見 - 米国”. www.afpbb.com (2007年4月13日). 2023年10月27日閲覧。
(17)^ “利用案内・情報 ≫ ホットニュース ≫ 2009-05-15 :: 国立科学博物館 National Museum of Nature and Science,Tokyo”. www.kahaku.go.jp. 2023年10月27日閲覧。
(18)^ “鳥と恐竜を結び付けたティラノサウルスのタンパク質”. natgeo.nikkeibp.co.jp. 2023年10月27日閲覧。
(19)^ Bath, University of. “New research shows 'juvenile' T. rex fossils are a distinct species of small tyrannosaur” (英語). phys.org. 2024年1月4日閲覧。
(20)^ Longrich, Nicholas R.; Saitta, Evan T. (2024年3月). "Taxonomic Status of Nanotyrannus lancensis (Dinosauria: Tyrannosauroidea)—A Distinct Taxon of Small-Bodied Tyrannosaur". Fossil Studies (英語). 2(1): 1–65. doi:10.3390/fossils2010001. ISSN 2813-6284。
(21)^ abpublished, Sascha Pare (2024年1月3日). “Nanotyrannus vs. T. rex saga continues: Controversial study 'doesn't settle the question at all'” (英語). livescience.com. 2024年1月4日閲覧。
(22)^ Voris, Jared T.; Therrien, Francois; Zelenitzky, Darla K.; Brown, Caleb M. (2020年). "A new tyrannosaurine (Theropoda:Tyrannosauridae) from the Campanian Foremost Formation of Alberta, Canada, provides insight into the evolution and biogeography of tyrannosaurids". Cretaceous Research. 110: 104388. doi:10.1016/j.cretres.2020.104388. S2CID 213838772。
(23)^ abcdefghijデイヴィッド・E・ファストヴスキー [ドイツ語版]、デイヴィッド・B・ウェイシャンペル [英語版]︵著︶﹃恐竜学入門 ─かたち・生態・絶滅─﹄藤原慎一・松本涼子︵訳︶、真鍋真監訳︵第2版版︶、東京都文京区千石3丁目36-7: 東京化学同人、2018年5月25日︵原著2015年1月31日︶、185–190頁。ISBN 978-4-8079-0856-1。NCID BB17901360。OCLC 904989280。全国書誌番号:22535495、ASIN 4807908561。
(24)^ Kent A Stevens (2006年). "Binocular vision in theropod dinosaurs". Journal of Vertebrate Paleontology. 26(2). doi:10.1671/0272-4634(2006)26[321:BVITD]2.0.CO;2。
(25)^ https://www.edu.city.kyoto.jp/science/about/0212tyranno.pdf
(26)^ Lawrence M. Witmer; Ryan C. Ridgely (2009年8月26日). "New Insights Into the Brain, Braincase, and Ear Region of Tyrannosaurs (Dinosauria, Theropoda), with Implications for Sensory Organization and Behavior". The Anatomical Record. 292 (9). doi:10.1002/ar.20983。
(27)^ Martin Kundrát; Xing Xu; Martina Hančová; Andrej Gajdoš; Yu Guo; Defeng Chen (2018年). "Evolutionary disparity in the endoneurocranial configuration between small and gigantic tyrannosauroids". Historical Biology. 32(5): 620–634. doi:10.1080/08912963.2018.1518442。
(28)^ Darla K Zelenitsky; François Therrien; Yoshitsugu Kobayashi (2008年10月28日). "Olfactory acuity in theropods: palaeobiological and evolutionary implications". Proceedings of the Royal Society B. 276 (1657). doi:10.1098/rspb.2008.1075。
(29)^ Thomas D. Carr; David J Varricchio; Jayc C Sedlmayr; Eric M Roberts; Jason R Moore (2017年). "A new tyrannosaur with evidence for anagenesis and crocodile-like facial sensory system". Scientific Reports. 7. doi:10.1038/srep44942。
(30)^ “ティラノサウルスはこんな顔だった、最新報告”. ナショナル・ジオグラフィック協会 (2017年4月4日). 2020年7月20日閲覧。
(31)^ S.V. Saveliev; V. R. Alifanov (2007年). "A new study of the brain of the predatory dinosaur Tarbosaurus bataar (Theropoda, Tyrannosauridae". Paleontological Journal. 41: 281–289. doi:10.1134/S0031030107030070。
(32)^ abcdeダレン・ナイシュ、ポール・バレット︵著︶﹃恐竜の教科書 最新研究で読み解く進化の謎﹄吉田三知世︵訳︶、小林快次・久保田克博・千葉謙太郎・田中康平監訳︵第1版版︶、大阪府大阪市中央区淡路町4-3-6: 創元社、2019年2月20日、141–147頁。ISBN 978-4-422-43028-7。
(33)^ 北村雄一︵著︶﹃21世紀こども百科 恐竜館﹄真鍋真︵監修︶、小学館、2007年7月11日、100–101頁。ISBN 978-4092212510。
(34)^ Phil R. Bell; Nicolás E. Campione; W. Scott Persons; Philip J. Currie; Peter L. Larson; Darren H. Tanke; Robert T. Bakker (2017年6月7日). "Tyrannosauroid integument reveals conflicting patterns of gigantism and feather evolution". Biology Letters. 13. doi:10.1098/rsbl.2017.0092。
(35)^ 川上和人﹃鳥類学者 無謀にも恐竜を語る (生物ミステリー)﹄技術評論社、2013年、76頁。ISBN 978-4774155654。
(36)^ abc鹿間時夫﹃古脊椎動物図鑑﹄朝倉書店、1979年、64頁。ISBN 978-4254162226。
(37)^ ab福田芳生﹃図解 恐竜はどんな生物だったか その素顔と生活をさぐる﹄講談社、1987年1月20日、87-88頁。ISBN 9784061326750。
(38)^ Cau, Andrea (13 dicembre 2023). “Theropoda: Tyrannosaurus era verde”. Theropoda. 2024年1月11日閲覧。
(39)^ Currie, P. J. (1998年). "Possible evidence of gregarious behavior in tyrannosaurids". Gaia. 15: 271–277. doi:10.7939/R3348GX03。
(40)^ Joseph E. Peterson; Karsen N. Daus (2019年3月4日). "Feeding traces attributable to juvenile Tyrannosaurus rex offer insight into ontogenetic dietary trends". PeerJ.
(41)^ abMichael Greshko (2020年2月1日). “成長期のティラノサウルス発見 新種論争に決着か”. Nikkei Style. 日本経済新聞社・日経BP. 2020年12月19日閲覧。
(42)^ “ティラノサウルス、これまでの想像より食欲旺盛=研究”. ロイター (2011年10月13日). 2020年12月19日閲覧。
(43)^ ab“Live fast, die young: How dinosaurs lived viciously short lives” (英語). www.sciencefocus.com. 2024年2月23日閲覧。
(44)^ Richard A.Thulborn (1982年1月21日). "Speeds and gaits of dinosaurs". Palaeogeography, Palaeoclimatology, Palaeoecology. 38(3–4): 227–256. doi:10.1016/0031-0182(82)90005-0。
(45)^ James O Farlow; Matt B Smith; John M Robinson (1995年). "Body mass, bone "strength indicator," and cursorial potential of Tyrannosaurus rex". Journal of Vertebrate Paleontology. 15(4): 713–725. doi:10.1080/02724634.1995.10011257。
(46)^ ab﹁ティラノサウルスは俊足? 生体力学で迫る恐竜の走り﹂﹃日本経済新聞﹄2019年7月27日。2020年7月20日閲覧。
(47)^ abc真鍋真、對比地孝亘、ダニエル・L・ブリンクマン、ヒシグシャフ・ツォクバートルイ・ユンナム、小林快次、ツォクトバートル・チンゾリグ、田中康平、今井拓哉、ダーラ・ゼレニスキー、フランソワ・テリエン、グレゴリー・ファンストン、石垣忍、西村智弘、佐藤たまき、小西卓哉、新村龍也、小原正顕、小松俊文、タイラー・R・ライソン、イアン・ミラー、フェルナンド・E・ノバス 著、坂田智佐子 編﹃恐竜博2019 THE DINOSAUR EXPO﹄NHK、NHKプロモーション、朝日新聞社、2019年、146-147頁。
(48)^ Patrick M O'Connor; Leon P A M Claessens (2005年). "Basic avian pulmonary design and flow-through ventilation in non-avian theropod dinosaurs". Nature. doi:10.1038/nature03716。
(49)^ Bill Sellers. “Tyrannosaurus Simulation”. 2005年12月25日時点のオリジナルよりアーカイブ。2020年7月20日閲覧。
(50)^ abSean D.Smith; W. Scott Persons IV; Lida Xingc (2016年). "A tyrannosaur trackway at Glenrock, Lance Formation (Maastrichtian), Wyoming". Cretaceous Research. 61: 1–4.
(51)^ “T. rex was a champion walker, super-efficient at lower speeds”. マギル大学 (2020年5月13日). 2020年7月19日閲覧。
(52)^ abcdBrian Switek (2015年11月11日). “T・レックスのメニュー拝見、ときには共食いも”. ナショナルジオグラフィック協会. 2020年12月19日閲覧。
(53)^ abL.M. Lambe (1917年1月1日). "The Cretaceous Theropodous dinosaur Gorgosaurus". Memoir 100. Geological Survey of Canada. doi:10.4095/101672。
(136)^ David A Krauss; J.M. Robinson (2013年). "The biomechanics of a plausible hunting strategy for Tyrannosaurus rex". Tyrannosaurid Paleobiology: 251–264.
(137)^ 土屋健﹃ティラノサウルスはすごい﹄小林快次︵監修︶、文藝春秋、2015年6月19日。生態の項
(138)^ Andrew A Farke; Ewan D. S. Wolff; Darren H. Tanke (2009年1月28日). "Evidence of combat in Triceratops". PLOS ONE. doi:10.1371/journal.pone.0004252。関連項目[編集]
- ティラノサウルスを題材とした事物(名称のみのものも含む)
- ティラノサウルス (小惑星)
- T・レックス - イギリスのグラムロックバンド。1968年にデビューした頃はティラノザウルス・レックスと名乗った。
- ティラノサウルスを主題とする作品一覧
外部リンク[編集]
- Acta Palaeontologica Polonica
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||















{kind=link}
| 典拠管理データベース: 国立図書館 |
|
|---|